Tillbaka på 20-talet. A. Lotka, och något senare, oberoende av honom, V. Volter, föreslog matematiska modeller som beskriver konjugerade fluktuationer i antalet rovdjur och bytesdjur.
Modellen består av två komponenter:
C – antal rovdjur; N – antal offer;
Låt oss anta att i frånvaro av rovdjur kommer bytespopulationen att växa exponentiellt: dN/dt = rN. Men bytesdjur förstörs av rovdjur i en takt som bestäms av frekvensen av möten mellan rovdjuret och bytet, och frekvensen av möten ökar när antalet rovdjur (C) och bytesdjur (N) ökar. Det exakta antalet bytesdjur som påträffas och framgångsrikt äts kommer att bero på effektiviteten med vilken rovdjuret hittar och fångar bytet, d.v.s. från a' - "sökeffektivitet" eller "attackfrekvens". Således kommer frekvensen av "framgångsrika" möten mellan rovdjuret och bytet och, därför, hastigheten för att äta offren att vara lika med a'СN och i allmänhet: dN/dt = rN - a'CN (1*).
I frånvaro av mat går enskilda rovdjur ner i vikt, svälter och dör. Låt oss anta att i den aktuella modellen kommer populationsstorleken för ett rovdjur i frånvaro av mat på grund av svält att minska exponentiellt: dC/dt = - qC, där q är dödlighet. Döden kompenseras genom att nya individer föds i en takt som i denna modell tros bero på två omständigheter:
1) matkonsumtionshastighet, a'CN;
2) effektiviteten (f) med vilken denna mat passerar in i rovdjurets avkomma.
Predatorns fertilitet är alltså lika med fa’CN och i allmänhet: dC/dt = fa’CN – qC (2*). Ekvationerna 1* och 2* utgör Lotka-Wolter-modellen. Egenskaperna för denna modell kan studeras, linjeisokliner kan konstrueras som motsvarar en konstant populationsstorlek och med hjälp av sådana isokliner kan beteendet hos interagerande rovdjur-bytespopulationer bestämmas.
I fallet med en bytespopulation: dN/dt = 0, rN = a'CN eller C = r/a'. Därför att r och a' = const, är isoklinen för offret den linje för vilken värdet på C är konstant:
Vid en låg täthet av rovdjur (C) ökar antalet byten (N) tvärtom, det minskar.
På liknande sätt för rovdjur (ekvation 2*) med dC/dt = 0, fa’CN = qC, eller N = q/fa’, dvs. En isoklin för ett rovdjur kommer att vara en linje längs vilken N är konstant: Vid höga bytestätheter ökar rovdjurets populationsstorlek och vid låga tätheter minskar den.
Deras antal genomgår obegränsade konjugerade fluktuationer. När antalet bytesdjur är stort ökar antalet predatorer, vilket leder till att rovdjurens tryck på bytespopulationen ökar och därmed minskar antalet. Denna minskning leder i sin tur till en begränsning av rovdjur i maten och en minskning av deras antal, vilket orsakar en försvagning av rovdjurstrycket och en ökning av antalet bytesdjur, vilket återigen leder till en ökning av populationen av rovdjur , etc.
Populationer utför samma svängningscykel på obestämd tid under lång tid tills någon yttre påverkan ändrar deras antal, varefter populationer utför nya cykler av obegränsade svängningar. Faktum är att miljön ständigt förändras, och befolkningen kommer ständigt att skifta till nya nivåer. För att de svängningscykler som en population gör ska vara regelbundna måste de vara stabila: om en yttre påverkan ändrar populationsnivån måste de tendera till den ursprungliga cykeln. Sådana cykler kallas stabila, limit-cykler.
Lotka-Wolter-modellen låter oss visa huvudtrenden i förhållandet rovdjur-byte, vilket uttrycks i förekomsten av fluktuationer i bytespopulationen, åtföljd av fluktuationer i rovdjurets population. Huvudmekanismen för sådana fluktuationer är tidsfördröjningen som är inneboende i sekvensen av tillstånd från ett högt antal bytesdjur till ett högt antal rovdjur, sedan till ett lågt antal bytesdjur och ett lågt antal rovdjur, till ett högt antal bytesdjur, etc.
5) BEFOLKNINGSSTRATEGIER FÖR PREDATOR OCH PRIMAT
Relationen "rovdjur-byte" representerar länkarna i processen för överföring av materia och energi från fytofager till zoofager eller från lägre ordningens rovdjur till rovdjur högre ordning. Förbi Baserat på arten av dessa relationer särskiljs tre typer av rovdjur:
A) samlare. Rovdjuret samlar små, ganska många mobila offer. Denna typ av predation är typisk för många fågelarter (plover, finkar, pipkärnor, etc.), som bara spenderar energi på att leta efter offer;
b) riktiga rovdjur. Rovdjuret förföljer och dödar bytet;
V) pastoralister. Dessa rovdjur använder byten upprepade gånger, till exempel gadflies eller hästflugor.
Strategin för att skaffa mat bland rovdjur syftar till att säkerställa energieffektivitet för näring: energiförbrukningen för att få mat bör vara mindre än den energi som erhålls under dess assimilering.
Riktiga rovdjurär uppdelade i
"skördare" som livnär sig på rikliga resurser (inklusive planktonfisk och till och med bardvalar), och "jägare" som letar efter mindre riklig mat. I sin tur
"Jägare" delas in i "bakhåll" som ligger och väntar på byten (till exempel gädda, hök, katt, mantisbagge), "sökare" (insektätande fåglar) och "förföljare". För den sistnämnda gruppen kräver sökandet efter mat inte stora energiutgifter, men mycket av den behövs för att ta bytet (lejon på savannerna) i besittning. Vissa rovdjur kan dock kombinera delar av strategin för olika jaktalternativ.
Som i förhållandet "fytofag-växt" observeras inte en situation i naturen där alla offer äts av rovdjur, vilket i slutändan leder till deras död. Ekologisk balans mellan rovdjur och bytesdjur upprätthålls av speciella mekanismer, vilket minskar risken för fullständig utrotning av offer. Så det kan offren:
Fly från ett rovdjur. I det här fallet, som ett resultat av anpassningar, ökar rörligheten för både offer och rovdjur, vilket är särskilt typiskt för stäppdjur som inte har någonstans att gömma sig för sina förföljare;
Skaffa skyddande färg (”låtsas” vara löv eller kvistar) eller omvänt, ljus färg, N.: röd, varnande rovdjur om den bittra smaken. Det är välkänt att harens färg ändras vid olika tider på året, vilket gör att den kan kamouflera sig i gräset på sommaren och mot bakgrund av vit snö på vintern. Adaptiva färgförändringar kan förekomma i olika stadier av ontogenesen: sälungar är vita (snöfärg) och vuxna är svarta (stenig kustfärg);
Fördela i grupper, vilket gör att leta efter och fånga dem mer energikrävande för rovdjuret;
Göm dig i skyddsrum;
Gå till aktiva försvarsåtgärder (växtätare med horn, taggiga fiskar), ibland led (myskoxar kan ta upp ett "allroundförsvar" från vargar etc.).
I sin tur utvecklar rovdjur inte bara förmågan att snabbt förfölja bytesdjur, utan också ett luktsinne, vilket gör att de kan bestämma bytesplatsen genom lukt. Många arter av rovdjur sliter isär hålorna för sina offer (rävar, vargar).
Samtidigt gör de själva allt för att undvika upptäckt av deras närvaro. Detta förklarar renheten hos små katter, som spenderar mycket tid på toaletten och begraver avföring för att eliminera lukter. Rovdjur bär "kamouflagerockar" (strimmor av gäddor och abborrar, vilket gör dem mindre märkbara i snår av makrofyter, ränder av tigrar, etc.).
Fullständigt skydd mot rovdjur av alla individer i populationer av bytesdjur förekommer inte heller, eftersom detta inte bara skulle leda till att svältande rovdjur dör, utan i slutändan till en katastrof för bytespopulationer. Samtidigt, i frånvaro eller minskning av populationstätheten av rovdjur, försämras bytespopulationens genpool (sjuka och gamla djur behålls) och på grund av en kraftig ökning av deras antal undergrävs födotillgången.
Av denna anledning observeras sällan effekten av beroendet av populationsstorlekarna för bytesdjur och rovdjur - en pulsering av bytespopulationens storlek, följt av en pulsering av rovdjurspopulationens storlek med viss fördröjning ("Lotka-Volterra-effekten"). .
Ett ganska stabilt förhållande etableras mellan biomassan hos rovdjur och bytesdjur. Således tillhandahåller R. Ricklefs data om att förhållandet mellan rovdjurs- och bytesbiomassa varierar från 1:150 till 1:300. I olika ekosystem i den tempererade zonen i USA finns det för en varg 300 små vitsvanshjortar (vikt 60 kg), 100 stora wapitihjortar (vikt 300 kg) eller 30 älgar (vikt 350). Samma mönster hittades på savanner.
Med intensiv exploatering av fytofaga populationer utesluter människor ofta rovdjur från ekosystemen (i Storbritannien, till exempel, finns rådjur och rådjur, men inga vargar; i konstgjorda reservoarer där karp och andra dammfiskar föds upp finns inga gäddor). I det här fallet utförs rovdjurets roll av personen själv, och tar bort en del av individerna i fytofagpopulationen.
En speciell variant av predation observeras i växter och svampar. I växtriket finns cirka 500 arter som är kapabla att fånga insekter och delvis smälta dem med hjälp av proteolytiska enzymer. Rovsvampar bildar fångstanordningar i form av små ovala eller sfäriska huvuden placerade på korta grenar av mycelet. Den vanligaste typen av fälla är dock adhesiva tredimensionella nätverk som består av ett stort antal ringar som bildas som ett resultat av förgrening av hyfer. Rovsvampar kan fånga ganska stora djur, som rundmaskar. Efter att masken trasslat in sig i hyferna växer de in i djurets kropp och fyller den snabbt.
1.Konstanta och gynnsamma nivåer av temperatur och luftfuktighet.
2. Överflöd av mat.
3. Skydd från negativa faktorer.
4.Aggressiv kemisk sammansättning livsmiljö (matsmältningsjuicer).
1. Närvaron av två livsmiljöer: den första ordningens miljö är värdorganismen, den andra ordningens miljö är den yttre miljön.
Izv. universitet "PND", vol. 19, nr 2, 2011
FENOMEN AV DEN MATEMATISKA MODELLEN AV LOTKA-VOLTERRA OCH LIKNANDE DEN*
DI. Trubetskov
Den matematiska modellen Lotka-Volterra (ofta kallad "rovdjur-byte"-modellen) är tillämpbar för att beskriva olika processer inom biologi, ekologi, medicin, social forskning, historia, radiofysik och andra vetenskaper.
Denna översikt, som till stor del är metodologisk till sin natur, undersöker varianter av denna modell och liknande i relation till analysen av ett antal natur- och sociala fenomen.
Följande modeller diskuteras: modeller för interaktion mellan föroreningar och miljö; modell för klasskamp; modell för ett klasslöst samhälle från jägare-samlare; modell för militära operationer; viral modell av infektionssjukdom; en modell för spridning av epidemier, inklusive en modell för datavirusinfektion; en modell av samspelet mellan kognitiva och/eller emotionella lägen i hjärnan.
Nyckelord: Lotka-Volterra matematisk modell, rovdjur-byte-system, analys av natur- och sociala fenomen.
"...När man löser rättsfall, som när man ställer en diagnos av en läkare eller under experiment utförda av en kemist, spelar erkännandet av typiska mönster en avgörande roll."
Bernhard Schlink. Lämna tillbaka**
Introduktionsbaserad miljöexempel
I sin kärna är Lotka-Volterra-modellen en matematisk beskrivning av den darwinistiska principen om kampen för tillvaron, som Charles Darwin anger i sin självbiografi enligt följande. ”Efter att ha studerat många omständigheter kom jag fram till att det finns en kamp för tillvaron. Det slog mig en gång att under liknande omständigheter skulle gynnsamma arter fortsätta att utvecklas
"St. Petersburg: Publishing group "ABC-Classics", 2010, s. 67
Vägen som Darwin gick beskrivs väl i boken. Det är märkligt att Darwin äntligen blev övertygad om sina slutsatser efter att ha läst boken "An Essay on the Law of Population" av Thomas Robert Malthus. Denna uppsats spelade en viktig roll i det sociala och politiska livet i Storbritannien på 1890-talet, eftersom det uppfattades som en rimlig förståelse av samhällets naturlagar. På 1930-talet undertryckte malthusianska doktriner regeringens politik. Låt oss påminna om Malthus argument, som är förvånansvärt enkla. Eftersom mänskligheten enligt Malthus har en naturlig tendens att öka sina arter kunde inte livsmedelsproduktionen ligga kvar på samma nivå.
"Även om det finns en ungefärlig balans, som författaren hävdade: ett antal människor dör som ett resultat naturliga faktorer(hunger eller sjukdom) eller mänskliga handlingar (krig, sexuell avhållsamhet eller syndiga handlingar som barnmord), detta är nödvändigt för den övergripande balansen i tillvaron,
Malthus gick vidare och argumenterade: ... detta öde (döden) är den svagaste, fattigaste och sjukaste medlemmen av samhället. Det är inte möjligt att undkomma det förberedda ödet, det vill säga Guds vilja själv, men författaren varnade för det "Om du ger allmosor till de fattiga, kommer människosläktet att öka, och det kommer att bli brist på mat."
Den viktorianska tiden verkade bekräfta att Malthus hade rätt. Det förekom matupplopp, diskussioner om fattiga lagar och offentliga motreaktioner mot majslagarna. År 1834 etablerade det viktorianska etablissemanget, som svar på ändringar av fattiglagen, arbetshus där människor arbetade för bröd och vatten som ett alternativ till kyrkliga välfärdssamhällen.
Darwin levde i denna värld, var dess representant och rörde sig i samma kretsar som Malthus. Han utvidgar Malthus idéer till allt levande: i naturen pågår en kamp för livet, en kamp för tillvaron, där den värsta och svagaste organismen dör först, och mer utvecklade former, friskare och mer anpassade, vinner. Det är dessa individer som föder avkomma. Organismer anpassar sig snabbare till miljöförhållanden om liknande fall av kamp upprepas med vissa intervall.
Låt oss börja vår ytterligare presentation (som en påminnelse) med modeller av interagerande populationer - de första matematiska modellerna av Lotka och Volterra. Vi kommer att följa presentationen av kapitel 5 i boken, eftersom det är lämpligt för ytterligare beskrivning av modeller för interaktion mellan föroreningar och miljö.
Författarna pekar först och främst på Volterras arbete och noterar att en liknande modell "föreslogs av Lotka något tidigare (Volterra genomförde en mycket mer komplett analys av detta system, Lotka försökte varje gång betona sin prioritet när det gäller att spela in denna modell) ”,
Lotka-Volterra-modellen beskriver samspelet mellan två arter - en population av rovdjur och en population av bytesdjur.
Låt N(t) vara antalet byten, P(t) vara antalet rovdjur vid tidpunkten t. Då har Lotka-Volterra-modellen formen:
AN - bNP, - = -dP + cNP, (1)
a, b, c, d är positiva konstanter.
Låt oss komma ihåg att ekvationssystemet (1) är baserat på följande antaganden:
I frånvaro av rovdjur förökar sig byten obegränsat enligt ekvationen dN/dt = aN, som ibland kallas Malthus-ekvationen;
Rovdjur i frånvaro av offer dör ut enligt ekvationen dP/dt = -dP;
Termerna som är proportionella mot produkten NP betraktas som omvandlingen av energin från en källa till energin från en annan (effekten av påverkan av populationen av rovdjur på populationen av rovdjur, det vill säga resultatet av deras möte, är att minska tillväxthastigheten dN/dt för antalet bytesdjur med mängden NP, proportionell mot antalet rovdjur).
Om du introducerar nya variabler
() N(t)d () P(t)b
n(x) = -v(x) = -^,
då kommer ekvationssystemet (1) att ta formen
N - nv, - = YV(n - 1), dx dx
Ekvationssystem (2) har två fixpunkter: (0, 0) - sadel och (1,1),
för vilka egenvärden Jacobian matriser A
- ^1.2 = ±iVУ,
i = l/- 1, det vill säga jämviktspositionen (1.1) är icke-hyperbolisk och det är omöjligt att dra en slutsats om dess stabilitet baserat på linjär analys.
Fasbanorna för ekvationssystemet (2), som är känt, är integralkurvorna för ekvationen
dv (n - 1)v dn Y (1 - v)n’
vems lösning
yn + v - ln nYv = H.
Det kan visas (se t.ex.) att nära (1, 1) nivålinjerna för funktionen H är slutna kurvor som motsvarar periodiska lösningar av ekvationssystemet (2) (Fig. 1). Konstanten H i uttryck (3) bestäms av initialförhållandena, till exempel n(0) = n0 och v(0) = vo.
Den matematiska modellen som beskrivs av ekvationssystemet (2) är strukturellt
är instabil, det vill säga en liten förändring på högersidan i den kan leda till en kvalitativ förändring av lösningens beteende. Detta är dess främsta nackdel. Dessutom är det därför det är svårt att prata om sammanträffandet av teori och experiment.
En av anledningarna till modellens strukturella instabilitet är att den är konservativ, det vill säga den har en första integral. Som bokens författare betonar, trots modellens angivna huvudsakliga nackdel, tillåter ekvationssystemet (1) en att dra icke-triviala slutsatser, bekräftade av många observationer. Speciellt formuleras Volterras princip: om i systemet "rovdjur-byte" som beskrivs av modell (1) utrotas båda arterna jämnt och i proportion till antalet individer, ökar det genomsnittliga antalet offer, och genomsnittet antalet rovdjur minskar.
Boken ger två exempel på förekomsten av denna effekt i naturen. Ett är det sociala faktum som ledde Volterra till sin teori om kampen för tillvaron: under första världskriget minskade fisket i Adriatiska havet kraftigt, vilket till biologernas förvåning ledde till en ökning av rovdjur och en minskning av antalet byten. Det andra exemplet är mindre känt. Enligt Volterras princip visar den dubbla karaktären av användningen av insektsmedel (insekticider) för att bevara grödor på fälten. Nästan alla sådana kemikalier verkar inte bara på skadedjur, utan också på deras naturliga fiender, vilket ofta leder till ett ökat antal skadedjur och en minskning av till exempel fåglar som livnär sig på dessa skadedjur. Således, enligt Volterras princip, i ett ekosystem av rovdjur och bytesdjur, är bytespopulationen mer känslig för processen med proportionell minskning av individer i populationen.
I terminologin av V.I. Arnold, modellen som beskrivs av ekvationssystemet (1) kallas "rigid". Förresten, ett avsnitt av boken av V.I. Arnold "Hårda" och "mjuka" matematiska modeller (M.: MCNO, 2000) kallas "Hårda modeller som en väg till felaktiga förutsägelser." Med en liten förändring i modellen blir det mjukt när små termer läggs till på högra sidorna av (1), med hänsyn till till exempel konkurrens mellan byten om mat eller rovdjur om byten (specifika typer av ekvationer ges Nedan). Sedan i allmän syn Ekvation (1) kan skrivas om enligt följande:
Beroende på små korrigeringar ef (P, N) och t.ex. (P, N) kan scenarierna som visas i fig. 2.
Tolkningsmässigt är figur 1 intressant. 2, b. Enligt Arnold, om P är banditer och N är arbetare, leder systemets utveckling antingen till en kraftig ökning av antalet banditer, eller till att de nästan helt utrotas, eftersom de har rånat arbetarna så mycket att det inte finns något kvar. att ta. Systemet faller så småningom in i en region med värden på P och N så stora eller så små att modellen blir otillämplig eftersom evolutionens lagar förändras.
Här är några mer komplicerade rovdjursbytesmodeller.
Systemet "rovdjur-byte" med hänsyn till intraspecifik konkurrens.
aN - bPN + ef (P, N)
dP + cPN + t.ex.(P,N), t.ex< 1.
Ris. 2. a - jämviktstillståndet är stabilt: för någon initiala förhållanden efter lång tid är samma sak etablerad; b - jämviktstillståndet B är instabilt: systemet "trampar"; c - i ett system med ett instabilt stationärt tillstånd A etableras ett periodiskt läge C över tiden (till skillnad från system (1) (se fig. 1), sker svängningar med en viss amplitud, oberoende av avvikelsens litenhet)
där K\,2 är den potentiella kapaciteten hos ekologiska system, som bestäms av den tillgängliga mängden resurser och motsvarar gränsvärdena för befolkningsantal. I dimensionslösa variabler tar detta ekvationssystem formen
där N(t) = (a/c)u(x), P(t) = av(x)/b, t Konkurrensmodell
at, y = d/a, a = a/(cK\), v = d/(aK2).
it1=1-Ш-eNN f=1-f)-hNN (5)
Här är n, r2, e, h positiva konstanter.
Analys av ekvationssystemet (5) visar: om n populationer är linjärt beroende av m resurser, och m< n, то по крайней мере одна из популяций вымирает.
Modell för ömsesidighet (symbios)
ui(ri + aiiui + ai2u2),
u2 (Г2 + a2iui + a22u2).
För att beskriva mutualism räcker det att kräva det
ai2 > 0, a2i > 0.
En detaljerad analys av ovanstående modeller finns till exempel i böckerna och.
I slutet av avsnittet, efter presentationen, betraktar vi den generaliserade Lotka-Volterra-modellen, beskriven av ekvationssystemet
A(u) - B(u,v), - = -D(v) + C (u,v), (7)
där u och v är antalet bytesdjur respektive rovdjur, A(u) är reproduktionsfunktionen för bytesdjur i frånvaro av rovdjur; D(v) - funktion av utrotning av rovdjur i frånvaro av bytesdjur; funktionen B(u, v) beskriver rovdjurens ätning av bytesdjur; C(u, v) är effektiviteten av byteskonsumtion av rovdjur. Låt oss lista möjliga ytterligare faktorer för relationer inom och mellan befolkningen.
Icke-linjärt beroende av reproduktionshastigheten för bytespopulationer av densitet vid låga densitetsvärden (avsaknad av tillräckligt antal parningspar): A(u) = au2/(N + u), där a och N är positiva konstanter.
Intraspecifik konkurrens av bytesdjur: A(u) = au(l - u/K).
Mättnad av rovdjur: Bi(u) = bu/(1 + au) - rovdjurets trofiska funktion.
Icke-linjär karaktär av predation av rovdjur: Bi(u) = bu2/(1 + au).
Konkurrens mellan rovdjur om byte: B2(v) = bv/(1 + |3v). I detta fall är B(u,v) = Bi(u)B2(v).
Konkurrens mellan rovdjur om resurser som skiljer sig från byten: C(v) = v/(1 + v/Ki).
Icke-linjär karaktär av beroendet av ett rovdjurs reproduktionshastighet på populationstäthet vid låga densitetsvärden: C2(v) = cv/(Nv + v), medan C(u,v) = C1(u)C2(v) och C1(u) = Bl(u), D2(v) = cv, det vill säga C(u,v) = = cv/(Nv + v) Bi(u).
1. Matematisk modell av föroreningars interaktion med miljön, tolkad som en "rovdjur-byte"-modell
Situationen ”föroreningar-natur” kan tolkas som specialfall"predator-prey"-modell, där naturen agerar som offer och föroreningar agerar som rovdjur. Huvudantagandet bakom modellen är att miljön aktivt absorberar och bearbetar föroreningar upp till en viss gräns.
Ur kvalitativa överväganden, i miljö-föroreningssystemet, är följande tre fundamentalt olika interaktionsscenarier möjliga.
1. Med små utsläpp av föroreningar återvinner miljön det helt (hållbar situation).
2. Med en ökning av utsläppen av föroreningar, beroende på yttre förhållanden och slumpmässiga orsaker, kan miljön vara i tillfredsställande skick, eller så kan den gå under (bistabil situation)*.
3. Slutligen motsvarar den tredje situationen en miljökatastrof - naturens fullständiga utrotning.
Låt oss anta att den allmänna bakgrunden till föroreningar och miljöns tillstånd kan karakteriseras av följande variabler: föroreningskoncentration P respektive biomassadensitet E. Om det finns en permanent föroreningskälla kan föroreningsprocessen beskrivas med ekvationen
där a är effekten av föroreningskällan per tidsenhet, b är koefficienten för naturlig förstörelse av föroreningar (sådan förstörelse kallas "död", eller
"En bistabil situation kan uppstå när, i närheten av vissa företag, vegetationstäcket bara finns kvar på vissa platser. Detta följer av observationer och kan på många sätt fungera som en försenad signal för att vidta akuta åtgärder för att rädda naturen.
naturlig spridning). Under initialvillkoret P(t = 0) = Po har lösningen till ekvation (8) formen:
P (t) = b+(Po - D e~bt’
det vill säga med tiden minskar koncentrationen av föroreningar naturligt.
Låt oss vidare anta att föroreningar är i ständig växelverkan med miljön, vilket har en renande effekt på föroreningarna. Vi kommer också att överväga att "miljö - föroreningar"-systemet är stängt. Sedan kan processen för interaktion med omgivningen beskrivas med följande ekvationssystem:
J- = a - bP- f (E, P), - = g(E) - h(E, P), (9)
där funktionen f (E, P) > 0 - beskriver absorption och bearbetning av miljöföroreningar; g(E) är en term som beskriver miljöns dynamik i frånvaro av föroreningar; h(E, P) är en funktion som beskriver föroreningars skadliga effekter på miljön.
Låt oss ta f (E, P) = cEP och h(E, P) = dEP som funktioner av interaktionen mellan föroreningar och vilda djur, där c och d är konstanta koefficienter.
Vi kommer att anta att i frånvaro av föroreningar kan miljöns beteende beskrivas med en logistisk ekvation, det vill säga g(E) = rE(1 - E/K), där r - konstant koefficient, och K - motsvarar det maximala värdet av E vid dE/dt = 0. Genom att ersätta uttrycken för f (E, P), h(E, P) och g(E) i ekvationssystemet (9), vi får
^ = a - bP-cEP, = rE (l -E) - dEP.
Övergång i ekvationssystemet (10) till dimensionslösa variabler
T = bt, a = -pr, uo
vi får den enklaste matematiska modellen av interaktionen mellan föroreningar och miljön i form av följande ekvationer:
A - u - uv, dt
v(u0 - u) - pv2.
Det är lätt att se att ekvationssystemet (10) är ett rovdjur-byte-system, där bytet är föroreningar (inte ett biologiskt objekt), och den biologiskt aktiva miljön är rovdjuret.
I ekvationssystemet (11) kan parameter a tolkas som den generaliserade effekten hos föroreningskällan; uo är den högsta tillåtna koncentrationen för ett givet ekosystem (om u > u0, dv/dt< 0, и природа вымирает); p - характеристика экосистемы - коэффициент внутривидовой конкуренции природы.
Jämviktspositionerna för ekvationssystemet (11), motsvarande villkoren du/dt = 0 och dv/dt = 0, är lätta att hitta från ekvationerna
a - u(l + v) = 0, -v(u - uo) - pv2 = 0.
Då Ai = (a, 0); A2 ((uo + p + Q)/2, (uo - p - Q)/(2p));
A3 ((uo + p - Q)/2, (uo - p + Q)/(2p)), Q = ^/(uo + p)2 - 4ap.
De andra och tredje jämviktspositionerna existerar om [(u + p)2-4aP]>0. Standard linjär analys av stabiliteten för de angivna jämviktspositionerna leder till ett parametriskt porträtt av ekvationssystemet (11), som visas i fig. 3. I fig. Figur 4 visar motsvarande fasporträtt av ekvationssystemet (11).
Om parametrarna tillhör region IV, tenderar alla banor med undantag av det stabila grenröret Ai att A3 som t ^ x (Fig. 4, a). Region II motsvarar fasporträttet i fig. 4, b. I region III finns en enda global atttraktor A1 (fig. 4, c).
Alltså den beskrivna matematisk modell motsvarar de tre kvalitativa situationer som beskrivs ovan.
I början av detta avsnitt angavs dock som huvudantagande att modellen innehåller ett tröskelvärde för föroreningar som naturen kan bearbeta. Denna effekt måste beaktas i den matematiska modellen.
Ris. 3. Parametriskt porträtt av ekvationssystem (11). För fast u0 är hela området med tillåtna parametrar uppdelat i fyra uppsättningar
För att göra detta föreslår boken att du använder en av de trofiska funktionerna som beskriver mättnadsprocessen, till exempel,
Sedan, vid en låg koncentration av föroreningar f (E, P) ~ cPE, vid en hög koncentration f (E, P) cE, vilket är mer realistiskt än det ursprungliga uttrycket.
Efter att ha reducerat ekvationssystemet (9) till en dimensionslös form, med hänsyn till relation (12) och omdesigneringar i, erhölls följande ekvationssystem:
A - u ---, - = v(uo - u) - pv , dt k + u dt
där k = A(d/b) > 0 beskriver graden av naturens påverkan på föroreningar: ju större dess värde, desto mindre är graden av absorption av levande natur och vice versa.
Boken anger också att i ekvationssystemet (13) är ett, två eller tre jämviktstillstånd av samma karaktär som för system (11) möjliga. Dessutom betonar författarna att a priori antaganden om valet av vissa funktionella beroenden i denna modell är mer verklighetsanpassade än de valda modellerna (11).
2. Matematisk modell för rening av avloppsvatten
Vi kommer att betrakta föroreningen som ett "byte" och biologiskt aktivt slam som ett "rovdjur". Processen med biokemisk oxidation av en förorening kommer att tolkas som att den "äts" av aktiverade slammikroorganismer.
Låt oss anta att det finns en konstant föroreningskälla, och aktivt slam kan behandla föroreningar till en viss gräns. Vi tror att förändringen i koncentrationen av aktivt slam i rent vatten minskar enligt en exponentiell lag.
Sedan kan dynamiken i avloppsvattenrening beskrivas med ekvationerna
dd _ = a - bD(P) - cf (P, E), - = -dS + eh(P, E),
där P (t) är koncentrationen av vattenföroreningar, E (t) är densiteten av aktiverad slambiomassa, D(P) är den avledningsfunktion som kännetecknar det naturliga förfallet av föroreningar; f (P, E) och h(P, E) - trofiska funktioner som kännetecknar processen för rening av föroreningar med biologiskt rent slam; a > 0 - föroreningskällans effekt; d > 0 är en konstant som kännetecknar minskningshastigheten för aktivt slam i rent vatten; c och e är positiva konstanter.
Låt oss anta att D(P) = P, och de trofiska funktionerna för "rovdjuret" och "bytet" är desamma och har formen:
f (P, E) = h(P, E) = , där r > 0 är en konstant.
Då har vi:
A - bP--------, -
Efter att ha introducerat dimensionslösa variabler m = at, P = ru(x), E = (rd/c) v(x), tar ekvationssystemet (14) formen
där a = a/(rd), b = b/d, y = e/d. Systemets fasta punkter (15) Ai(ui,vi) =
= (a/fr 0) och A2(u2, v2) = (1/(Y - 1), Hy - 1) - p]y/(y - 1)).
Standard linjär analys visar att punkten Ai(ui, vi) är en sadelpunkt vid< a(Y - 1) и y >0 och en stabil nod för β > a(Y - 1).
Om du ökar värdet på parametern a vid fasta värden i och (Y - 1), vilket motsvarar en ökning av föroreningskällans intensitet, så uppstår ett stabilt jämviktstillstånd A^(u2,v2). Om parametern a är liten (föroreningsintensiteten är låg), tenderar massan av biologiskt aktivt slam till noll. Eftersom föroreningen är mat för biologiskt aktivt slam, försvinner det biologiskt aktiva slammet om intensiteten av föroreningen är otillräcklig.
Det är rimligt att anta att det finns ett visst tröskelvärde för föroreningskoncentrationen P*, över vilket reningskapaciteten av slam minskar, och att beakta artens trofiska funktioner
f (P, E) = EPe-rP, r = const > 0.
I detta fall är föroreningströskelvärdet P* = 1/r. I verkligheten finns det ytterligare en komponent i den beskrivna processen - luftningsprocessen - mättnad av slam med syre, vilket avsevärt ökar förmågan hos biologiskt aktivt slam att bearbeta föroreningen. Med hänsyn till ovanstående kommer den matematiska modellen för rengöring att ta formen:
a - bP - cf (P, E),
dE + ef (P, E)Q(t),
där a, b, c,d,e = const > 0. Här är Q(t) syrekoncentrationen, och R> 0 bestämmer mängden syre som strömmar in i systemet per tidsenhet.
Analysen av ekvationssystemet (16) utförd av författarna till boken under antagandet att f (P, E) = Pg(E), där g(E) är en jämn monotont ökande icke-negativ funktion (och g(0) = 0), visar att när Med tillräckligt nära värden för parametern R, som bestämmer mängden syre som kommer in i systemet, är det alltid möjligt att uppnå systemstabilitet.
3. Modellerande klasskamp
Låt oss skriva ekvationerna för "rovdjur-byte"-modellen i form av följande system:
där x, y är populationstalen, x1 och y1 är deras stationära värden, a och b är konstanter.
Denna modell används för att beskriva små stadsområden. I detta fall betyder variabeln x i ekvationssystemet (17) markanvändningsarean, y - markränta, a, b, xi, yi - några parametrar.
En intressant tillämpning av modellen för att beskriva klasskamp ges av Goodwin. Låt oss överväga två typer av medborgare: arbetare och kapitalister.
Arbetare spenderar all sin inkomst wL på konsumtion, kapitalister ackumulerar sin inkomst Y - wL, där Y är produktionsproduktion. Priset på konsumtionsvaror är normaliserat till ett.
Låt K betyda kapital, a = ао egt = Y/L - arbetsproduktivitet, ökar i konstant takt d, k = K/Y - koefficienten för produktionens kapitalintensitet, N = Noent - utbudet på arbetsmarknaden, vilket ökar med tillväxttakten n Andelen arbetskostnader i förhållande till nationalinkomsten är wL/Y = w/a. Därför är kapitalisternas vinstandel (1 - w/a). Eftersom sparande kan definieras som S = Y - wL = (1 - w/a)Y, är investeringsandelen dK/dt = S = (1 - w/a)Y eller (dK/dt)(1/K) = (1-w/a)(Y/K); I det här fallet försummar vi pensioneringen av kapital.
Med ett konstant värde på kapitalintensiteten k = K/Y får vi dk/dt = 0 och
dY dL 1 dt Y dt L
Dessutom, från (18), med hänsyn till det faktum att dK/dt = (1 - w/a)L, följer det
dY 1 dK 1 / w \ 1
dt Y dt K V a) k-
Från relationer (19) och (20) finner vi slutligen:
Låt oss introducera nya variabler: y = w/a - andel av arbetskostnaderna; x = L/N - beläggningsgrad.
Efter enkla transformationer, med hänsyn till (21) får vi
dx - = x dt 1 - y k (d + p)
dt =у w dt a dt
Wf (x), lim f (x) dt
limf(x)< 0,
En linjär approximation av f (x) i formen (dw/dt)(l/w) = -r + bx förvandlar (23) till följande ekvation:
R + px - g. (24)
Kombinera (22) och ekv.
(24) kommer vi fram till Goodwin-modellen, som beskrivs av systemet
y[-(g + r) + px).
Det är uppenbart att ekvationssystemet (25) är ekvivalent med ekvationssystemet (17), därför är alla allmänna slutsatser som är giltiga för system (17) även sanna för Goodwin-modellen.
Notera att Goodwins modell, som tar hänsyn till växelverkan mellan sysselsättningsnivån och den juridiskt fastställda andelen bidrag till lönerna, påminner mycket om klassiska modeller för politisk ekonomi (den kallas ibland neomarxistisk). Modellen uppmärksammar återigen verk av klassiska ekonomer som Ricardo, Smith och Marx. Tyvärr är modellen strukturellt instabil.
Så, "rovdjur"-"byte"-modellen kan användas för att modellera fenomenet av förhållandet mellan urban markhyra och intensiteten av markanvändning, arbetslöshet och dynamiken i ekonomisk tillväxt.
4. "rovdjur-byte"-modellen för att beskriva ett klasslöst samhälle - ett samhälle från jägare-samlare
Denna era kännetecknas av låg befolkningstäthet och kommunal (stam) organisation av det mänskliga samhället. Människan var helt beroende av naturen, det pågick en ständig kamp för överlevnad och den huvudsakliga resursen som utvanns var mat. Människan var en del av ekosystemet och skilde sig från andra skolrovdjur, i huvudsak bara i förmågan att använda eld och primitiva verktyg för sina egna behov. Förändringen i populationsstorlek i ett visst område bestämdes huvudsakligen av resursbasens tillstånd och kan beskrivas med modellen "rovdjur-byte" - ett ekvationssystem (1). I dessa ekvationer är N nu populationsstorleken för jagade föremål ("offer"); P är antalet forntida jägare ("rovdjur") i det aktuella området; cNP - födelsehastighet för "rovdjur"; dP - dödlighet för "rovdjur"; aN är dödsfrekvensen för "offer" (man tror att deras mattillgång är obegränsad); bNP är dödsfrekvensen för "byten" på grund av att de utrotas av "rovdjur" (man tror att detta är den enda orsaken till dödligheten för "byten". Lösningar på dessa ekvationer analyserades i inledningen). Eftersom jägare-samlare konsumerade det som producerades i naturen utan deras deltagande, bestämdes populationsstorleken av yttre förhållanden som de inte kunde påverka (i modellen fluktuerar antalet jägare-samlare runt medelvärdet
Po = a/b. Observera att detta förklarar den relativa stabiliteten hos jordens befolkning under hela stenåldern fram till den neolitiska eran.
När människan tämjde djur och lärde sig att odla grödor förvandlades hon från en jägare-samlare till en pastoralist och bonde. Således ökade koefficienten a och jämviktsantalet personer avsevärt, eftersom värdet på koefficienten b, som kännetecknar konsumtionen, höll sig ungefär på samma nivå.
5. Liknande ideologi av människor för att beskriva militära handlingar
I boken föreslog Frederick Lanchester - en engelsk polymat och ingenjör - en enkel modell av kampen mellan två motståndare (säg två arméer), som bär hans namn. Observera att samma modell föreslogs av tsararméofficeren M.O. Osipov 1915. Därför är det rättvist att kalla denna modell Lanchester-Osipov-modellen. Vidare i presentationen kommer vi att följa arbetet. I denna modell beskrivs systemets tillstånd av punkten (x, y) i den positiva kvadranten av planet. X- och y-koordinaterna för denna punkt är antalet motsatta arméer. Modellekvationerna är följande:
Av, - = -ax. (26)
Här är a och b kraften hos armé x respektive armé y vapen. Med andra ord, varje soldat av armé x dödar en soldat av armé y per tidsenhet (och följaktligen dödar varje soldat av armé y b soldater av armé x). Således ges den totala effektiviteten av alla skott av armé x av yxa och för armé y av by. Arbetet förutsätter att den kontinuerliga approximationen är tillräckligt bra, och förändringshastigheten i antalet trupper är proportionell mot effektiviteten av fiendens skott. "Hårt", enligt V.I. Arnold, modell (26) medger en exakt lösning i formen:
ax2 - bу2 = konst. (27)
Förändringen i antalet arméer x och y sker längs hyperbeln, ges av ekvationen(27) (Fig. 5). Vilken överdrift kriget kommer att följa beror på utgångspunkten. Hyperbolerna är åtskilda av den räta linjen l/ax = \[bу. Om startpunkten ligger ovanför denna linje, går hyperbeln till y-axeln. Det betyder att under kriget minskar storleken på armé x till noll på en begränsad tid. Armén vinner, fienden förstörs. Om startpunkten ligger under linjen vinner armé x. I en rak linje, som Arnold skriver, slutar kriget "till allas belåtenhet" med utrotningen av båda arméerna. Men detta tar oändligt lång tid: konflikten fortsätter att glöda när båda motståndarna redan är utmattade. Av relation (27) följer att på den räta linjen ^fax = -\[bу. Sedan tar den första ekvationen av systemet (26) formen: dx/dt =
Y/abx. Dess lösning är x = Ce~^a>t, där
С = och det är uppenbart att x > 0 vid t > Ж. 5. "Hård" modell av krig
Ris. 6. "Mjuk" modell av krig
Av modell (26) följer att för att bekämpa en fiende dubbelt så många, är det nödvändigt att ha fyra gånger kraftfullare vapen, med tre gånger så många - nio gånger kraftfullare, etc. Detta indikeras av kvadratrötterna i linjens ekvation.
Det är tydligt att den resulterande "kannibalistiska" modellen, i Arnolds terminologi, är mycket idealiserad, och det är farligt att tillämpa den på en verklig situation. Vad kommer att förändras om
Detta ekvationssystem kan inte längre lösas explicit. Men modellen blir "mjuk" på grund av förändringar i a(x, y) och b(x, y). Kurvorna på planet (x, y) är inte längre hyperboler och de är inte åtskilda av en rät linje (fig. 6). Men den huvudsakliga kvalitativa slutsatsen förblir densamma: situationerna "x vinner" och "y vinner" är åtskilda neutral linje"båda arméerna förstör varandra på oändlig tid."
6. Den enklaste virala modellen av en infektionssjukdom
1974 föreslog G. Bell den enklaste modellen av immunsvaret, där interaktionen mellan antigen och antikropp beskrivs i termer av "rovdjur-byte".
Låt oss presentera en version av G. Bells teori, baserad på den mer allmänna modellen av G.I. Marchuk.
Huvudfaktorerna i modellen för infektionssjukdomar är följande:
Koncentration av patogena multiplicerande antigener V(t);
Antikroppskoncentration F(t); Antikroppar betyder substrat för immunsystemet.
system som neutraliserar antigener (immunoglobuliner, cellreceptorer);
Koncentrationen av plasmaceller C - bärare och producenter av antikroppar - antas vara konstant;
Graden av målorganskada tas inte med i beräkningen.
Ekvationen som beskriver förändringen av antalet antigener (främmande celler som penetrerar kroppen) har formen
W = eV - YFV (28)
Den första termen i ekvation (28) är tillväxthastigheten för antigener på grund av reproduktion; c - antigen multiplikationskoefficient; termen yFV beskriver antalet antigener neutraliserade av F-antikroppar per tidsenhet; y är en koefficient associerad med sannolikheten för neutralisering av antigenet av antikroppar när de möter dem.
För att få den andra ekvationen, låt oss beräkna balansen av antalet antikroppar som reagerar med antigenet. Kommer att ha
dF = pCdt - nyFVdt - c/Fdt. (29)
Den första termen på höger sida av ekvation (29) beskriver genereringen av antikroppar av plasmaceller under tiden dt; p är hastigheten för antikroppsproduktion av plasmacellen. Den andra termen (nyFVdt) beskriver minskningen av antalet antikroppar på grund av kommunikation med antigener, och det finns n antikroppar för att neutralisera ett antigen. Den tredje termen (ifFdt) beskriver minskningen av antikroppspopulationen på grund av åldrande; if är en koefficient omvänt proportionell mot sönderfallstiden för antikroppar. Dividera ekvation (29) med dt får vi
RS - (om + nyV)F. (trettio)
Om vid t< to V(t) = 0, то начальные условия имеют вид
V (to) = Vo, F (to) = Fo. (31)
Låt in det som följer låt till = 0.
Låt oss överväga två extrema fall av sjukdomsdynamik. Låt oss anta att kroppen inte producerar antikroppar med denna specificitet, det vill säga F(t) = Fo = 0 för alla t > 0 och p = 0. Av (28) följer det att
- = |3V och V (t) = Voeet, (32)
där Vo är infektionsdosen (initial koncentration av antigener) vid t = 0. Uppenbarligen motsvarar denna lösning sjukdomsförloppet med dödlig utgång, eftersom det inte finns några faktorer som kompenserar för tillväxten av antigener. Det andra begränsningsfallet: de antikroppar som finns i kroppen (specifika för ett givet antigen) är tillräckliga för att förstöra alla antigener som har kommit in i kroppen, utan att utlösa mekanismen för antikroppsbildning. I detta fall, i ekvation (28) i ^ yF. Om man antar att infektionsdosen Vo är liten kan värdet på F anses konstant, bestämt av den normala nivån av antikroppar F*. Då har ekvation (28) formen
F = (v - YF*> V
och hans lösning är skriven så här:
V = Voe-(YF *-e)t.
I begränsningsfallet när B = 0
V = Voe-YF h, (33)
vilket motsvarar en hög immunologisk barriär.
Lösningar (32) och (33) presenteras i fig. 7.
För givna värden på modellkoefficienterna och initiala förhållanden är det naturligt att hela familjen av olika sjukdomsdynamik passar in i det skuggade området i fig. 7.
begränsas av lösningar för två begränsande fall
7. Liknande mönster för epidemispridning
Betrakta Kermack-McKendrick SIR-modellen, förutsatt att individer i en population kan vara i tre olika tillstånd:
S (t) - friska individer som är i riskzonen och kan drabbas av en infektion;
I(t) - smittade sjukdomsbärare;
R(t) - de som har återhämtat sig och slutat sprida sjukdomen (denna kategori inkluderar till exempel de som har förvärvat immunitet eller till och med de som har dött).
Den typiska utvecklingen av en individ i en population beskrivs av följande diagram:
Låt oss bygga en fenomenologisk modell som motsvarar diagrammet med hjälp av följande antaganden:
1) populationen är sluten och har en konstant storlek N;
2) tidsperioden då den sjuke förblir farlig för andra fördelas enligt en exponentiell lag med ett genomsnitt på 1/år;
3) kontakter är slumpmässiga och lika möjliga (likformig blandning).
Under dessa antaganden kommer vi att ha:
där X är den så kallade infektionsstyrkan, som i detta fall är lika med produkten av antalet kontakter per tidsenhet som gjorts av en frisk person (vi betecknar detta nummer som cN), sannolikheten att överföra infektionen till honom vid kontakt p och sannolikheten att träffa en sjuk person I(t) /N. Låt oss beteckna med = cp. Då får vi X = |3! och vi kommer fram till standardformen för ekvationerna:
w dt = S-■<’" ~dt = -о* d + d. + dt = 0и
S (t) + I (t) + R(t) = N.
Den tredje ekvationen i det reducerade systemet är redundant och eftersom dS/dt< 0, то естественно считать фазовым пространством треугольник S(t) + I(t) < N.
Av de första och andra ekvationerna av systemet (34) följer det
dL =(М-Т^ = -1+ о о = Y I = о dS eSI S’ о в, I =
Med hänsyn till initialvillkoren S(0) = Så, I(0) = I0, hittar vi den första integralen I + S - o ln S = I0 + S0 - o ln S0, med hjälp av vilken vi kan konstruera fas kurvor på planet (S, I) (fig. 8). Kurvorna som visas i fig. 8, b, sammanfaller väl med uppgifter om pestepidemin i Bombay 1906.
Boken föreslår att man ska använda modellen som diskuterats ovan, med viss modifiering, för att analysera datavirusinfektioner. Problemformuleringen är som följer.
Låt S vara antalet datorer som är infekterade med ett virus, jag är den del av datorer som är infekterade med ett virus och inte har antivirusprogram, och R är den del av datorer som har korrekt antivirusskydd ( immunitet). Sedan har modellekvationerna formen (se ekvationssystem (34)):
^ = -aSI, ^ = aSI - bI, ^ = bI. dt dt dt
Huvudidén med att modifiera denna klassiska epidemimodell är att introducera ett element av cyklicitet, det vill säga ett element för att reglera tillägget av nya datorer som är mottagliga för epidemin. Vid varje ny tidsperiod beräknas antalet infekterade datorer baserat på antalet infekterade datorer som finns kvar från föregående ögonblick, och antalet datorer som exponerats för epidemin spelas upp som en slumpmässig variabel enligt Poissons lag med fördelningen pm = (Xm/m\)e-x, m = 1,2, ...
Lösningen på problemet är inte given. Författarna föreslår följande lösning. Konstruera en graf av funktionen I(t) genom att numeriskt lösa ovanstående ekvationssystem med initialvillkoren 5(0) = 100, I(0) = 1, R(0) = 0, X = 10, a = 0,23 b = 0,5. Det föreslås att lägga till nya element vid tk = k, k = 1, 2, ...100.
Kanske kommer några av läsarna att vara intresserade av lösningen på detta problem och dess många alternativ.
8. Generaliserad Lotka-Volterra modell för beskrivning
interaktioner mellan kognitiva och/eller emotionella lägen i hjärnan
Enligt arbetet ska storskaliga kognitiva mönster (lägen eller representationer som observeras i ett experiment) i hjärnans funktionsläge undertrycka varandra, vilket naturligtvis bör inträffa sekventiellt över tiden. Den arbetande hjärnan uppvisar med andra ord kognitiv och emotionell aktivitet i form av en kedja av kombinationer av funktionssätt som avlöser varandra över tid, och dessa kombinationer i sig bestäms av typen av mental aktivitet.
Enligt författarna är "... konkurrens om ändliga resurser en av de huvudsakliga operativa krafterna som styr hjärnprocesser. Dessa resurser är energi (syre och glukos) och information (uppmärksamhet och minne). Det är viktigt att informationsresurser är en produkt av själva hjärnans aktivitet, det vill säga de kan påverka känslor och kognitiva funktioner på olika sätt (till exempel genom uppmärksamhetshantering) och bör ingå i den generella dynamiska modellen som definierar arbetsyta. ...Moder som representerar känslor och kognitiva funktioner på arbetsplatsen utbyter information ömsesidigt och tävlar om energi och informationsresurser."
funktionellt bestäms av samspelet mellan olika hjärnsubsystem, deras konkurrens och synkronisering (koordination) i tid." De ovan nämnda processerna för konkurrens mellan kognitiva och emotionella lägen med varandra, såväl som emotionella och kognitiva lägen med varandra, föreslås beskrivas i arbetet med ekvationssystem av Lotka-Volterra-typ i följande generaliserade form :
Vi(E) -^2 Wi,j(E)xi(t)
Här kännetecknar Xi(t) aktiviteten hos det i:te läget (storleken på den i:te populationen inom ekologi); n är antalet interagerande moder (populationer); u(E) - information som kommer in i systemet eller tillgängliga resurser; f^- (E) - element i interaktionsmatrisen; n(t) är det multiplikativa bruset som finns i systemet; t är den karakteristiska tiden som bestämmer processen.
Om vi tar hänsyn till särdragen hos de resurser som kognitiva och emotionella lägen konkurrerar om, bör det finnas tre delsystem av ekvationer (36): emotionella lägen, kognitiva lägen och resurser (uppmärksamhet, minne, energi). Uppmärksamhetens roll är särskilt viktig. I artikeln, för enkelhetens skull, antas det ... att beskrivningen av konkurrensen om uppmärksamhet mellan känslor och kognitiva funktioner inte kräver specifikation av individuella lägen, och därför kan vi begränsa oss till att överväga konkurrensen om uppmärksamhet "i allmänhet ” av känslor B = ^П = 1 Bi och ”i allmänhet » kognitiva lägen A = ^П=1 Ai. Då kan de grundläggande ekvationerna skrivas som:
Oi(I,B,D)Ra -^2 Pij(D)Aj(t)
Zi(S, A,D)Rb -£ lij (D)Bj(t)
Ai(t)n(t), i = 1,...N,
Bi(t)n(t), i = 1, ...M,
RA(t) A - (RA(t) + Fl(1, D)Rb(t)) = Rb(t) B - (Rb(t) + Fb(S, D)RA(t))
I dessa ekvationer är Ai och Bi icke-negativa variabler som motsvarar kognitiva och emotionella moder, vars totala intensitet betecknas med A respektive B; I - information från omvärlden; S - karakteristisk för känslomässig påverkan från omvärlden (vid negativa känslor - stress); D - nivå av mediciner som tagits; ta och tv är karakteristiska tider av medveten och känslomässig aktivitet; n(t) - multiplikativt brus; koefficienterna Fa och Fv bestämmer intensiteten i kampen för uppmärksamhet från känslor och tänkande. Parametrarna Oi och Zis beroende av A respektive B beskriver det direkta inflytandet av kognitiva processers aktivitet på känslor och känslor på tänkande. I artikeln
en datoranalys av ekvationerna ovan presenteras i närvaro av en extern stimulans S(t), som kännetecknar stressnivån. Samspelet mellan humör och tänkande diskuteras också. Interaktionen mellan tre komponenter i hjärnans aktivitet analyseras: beteende, känslomässigt minne och kognitiv aktivitet i frånvaro av yttre påverkan.
Istället för en slutsats
I den underbara boken av Yu.I. Neymark har sådana ord.
"Vad är en modell? Ordet "modell" kommer från det italienska språket och betyder "kopia, prov, prototyp", genom att studera vilket vi lär känna originalet. Detta betyder att en matematisk modell är en prototyp av någon del av världen - ett objekt, system, enhet, maskin, process, enhet, genom att studera vilken vi förstår denna del av världen...”
Om modellen är standard (se epigraf) eller, som de säger, standard, kan du med dess hjälp förstå inte en, utan olika delar av världen. Den matematiska modellen Lotka-Volterra är just en sådan modell. Naturligtvis täcker den presenterade recensionen endast en del av dess möjliga tillämpningar och gör inte anspråk på att vara universell.
Bibliografi
1. Kap. Darvin. Självbiografi. 1958. S. 120.
2. Brun Janet. Charles Darwin. Arternas ursprung / Ser. "10 böcker som förändrade världen." M.: AST: ast., 2009. 220 sid.
3. Malthus T.R. En analys av principen om befolkning, eftersom det påverkar den framtida förbättringen av samhället. 1798.
http://www.faculty.rsu.edu/felwell/Theorists/Malthus/essay2.htm
4. Lotka A. Elements of Physical Biology. Baltimore, 1925. Omtryckt av Dover 1956 som Elements of Mathematical Biology.
5. Volterra V. Matematisk teori om kampen för tillvaron / Övers. från franska M.: Nauka, Huvudredaktionen för fysisk och matematisk litteratur, 1976. 288 sid.
6. Bratus A.S., Novozhilov A.S., Platonov A.P. Dynamiska system och modeller av biologi. M.: FIZMATLIT, 2010. 400 sid.
7. Bazykin A.D. Icke-linjär dynamik hos interagerande populationer. Moskva-Izhevsk: Institutet för datorforskning, 2003. 368 s.
8. Arnold V.I. "Hårda" och "mjuka" modeller // Nature. 1998. Nr 4. P. 3.
9. Bratus A.S., Meshcherin A.S., Novozhilov A.S. Matematiska modeller för interaktion mellan föroreningar och miljö // Bulletin of Moscow State University. Ser. Beräkningsmatematik och cybernetik. 2001. T. 6. S. 140.
10. Zang W.-B. Synergetisk ekonomi. Tid och förändringar i icke-linjär ekonomisk teori / Transl. från engelska M.: Mir, 1999. 335 sid.
11. Goodwin R.M. En tillväxtmodell // Socialism och tillväxt. Cambridge: University Press, 1967.
12. Malkov S.Yu. Social självorganisering och historisk process. Kapitel 2. M.: URSS, 2009.
13. Lanchester F W. Flygplan i krigseld: Nedgången av den fjärde armen. London, konstapel, 1916.
14. Osipov M.O. Om påverkan av antalet styrkor som går in i strid på deras förluster // Militär insamling, juni-oktober, 1915.
15. Bell G. Prey - rovdjursekvationer som simulerar immunsvar // Math. Biosci. 1973. Nr 16. S. 291.
16. Marchuk G.I. Matematiska modeller inom immunologi och medicin. Kapitel 2. M.: Nauka, 1985.
17. Kermack W.O. och Mc.Kendrick A.G. Bidrag till den matematiska teorin om epidemier // Proceedings of Royal Statistical Society A. 1927. Vol. 115. S. 700.
18. Rabinovich M.I., Muezinolu M.K. Icke-linjär dynamik i hjärnan: känslor och intellektuell aktivitet // UFN. 2010. Nr 4.
19. Neimark Yu.I. Matematiska modeller inom naturvetenskap och teknik. N. Novgorod: Nizhny Novgorod University Publishing House, 2004.
Saratov State Mottagen 2011-11-02
Universitet uppkallat efter N.G. Chernyshevsky
FENOMEN LOTKA-VOLTERRA MATEMATISK MODELL
OCH LIKNANDE MODELLER
Lotka-Volterra matematisk modell (ofta kallad "predator-prey" modell) är tillämpbar för olika processbeskrivningar inom biologi, ekologi, medicin, i sociologiska undersökningar, i historia, radiofysik, etc.
Varianter av denna modell övervägs metodologiskt i denna recension.
Följande modeller observeras: modell av kontaminering eller annan smutsig interaktion med omgivningar; modell för klasskamp; modell av klasslöst samhälle - epock av jägare-samlare; militär operationsmodell; modell av virusinfektionssjukdomar; modell av epidemi spridning, så virus av datorer infektion sprider sig; modell av kognitiva och (eller) känslor cerebrala lägen.
Nyckelord: Lotka-Volterra matematisk modell, "rovdjur-byte"-system, analys av natur- eller samhällsfenomen.
Trubetskov Dmitry Ivanovich - född i Saratov (1938). Utexaminerad från fakulteten för fysik vid Saratov State University (1960). Han disputerade för den vetenskapliga examen som kandidat (1965) och doktor i fysikaliska och matematiska vetenskaper vid SSU (1978) inom området radiofysik. Chef för avdelningen för elektronik, svängningar och vågor, fakulteten för icke-linjära processer, SSU, professor, korresponderande medlem av Ryska vetenskapsakademin, hedrad vetenskapsman i Ryska federationen, pristagare av presidentens pris i utbildning. Vetenskaplig chef för Lyceum of Applied Sciences och fakulteten för icke-linjära processer vid SSU. Område av vetenskapliga intressen: vakuumelektronik och mikrovågsmikroelektronik, teori om oscillationer och vågor, icke-linjär dynamik, vetenskapshistoria. Författare till mer än tjugo läroböcker och monografier, samt mer än tvåhundra artiklar i tidskrifter.
410012, Saratov, st. Astrakhanskaya, 83
Saratov State University uppkallad efter. N.G. Chernyshevsky E-post: [e-postskyddad]
Modeller för interaktion av två typer
Volterras hypoteser. Analogier med kemisk kinetik. Volterra modeller av interaktioner. Klassificering av interaktionstyper Konkurrens. Rovdjur-byte. Generaliserade modeller av artinteraktioner . Kolmogorov modell. MacArthurs modell för interaktion mellan två insektsarter. Parametrisk och fasporträtt av Bazykin-systemet.
Grundaren av den moderna matematiska teorin om populationer anses med rätta vara den italienske matematikern Vito Volterra, som utvecklade den matematiska teorin om biologiska samhällen, vars apparat är differential- och integro-differentialekvationer.(Vito Volterra. Lecons sur la Theorie Mathematique de la Lutte pour la Vie. Paris1931). Under efterföljande decennier utvecklades befolkningsdynamiken huvudsakligen i linje med de idéer som uttrycks i denna bok. Den ryska översättningen av Volterras bok publicerades 1976 under titeln: "The Mathematical Theory of the Struggle for Existence" med ett efterord av Yu.M. Svirezhev, som undersöker historien om utvecklingen av matematisk ekologi under perioden 1931–1976.
Volterras bok är skriven på det sätt som böcker om matematik är skrivna. Den formulerar först några antaganden om de matematiska objekt som är tänkta att studeras, och genomför sedan en matematisk studie av egenskaperna hos dessa objekt.
Systemen som studerats av Volterra består av två eller flera typer. I vissa fall övervägs tillgången på mat som används. Ekvationerna som beskriver interaktionen mellan dessa typer är baserade på följande begrepp.
Volterras hypoteser
1. Livsmedel finns antingen tillgängligt i obegränsade mängder eller också är dess utbud strikt reglerat över tiden.
2. Individer av varje art dör ut på ett sådant sätt att en konstant andel existerande individer dör per tidsenhet.
3. Rovdjur äter offer, och per tidsenhet är antalet uppätna offer alltid proportionellt mot sannolikheten att träffa individer av dessa två arter, d.v.s. produkten av antalet rovdjur och antalet byten.
4. Om det finns föda i begränsade mängder och flera arter som kan konsumera den, så är andelen föda som konsumeras av en art per tidsenhet proportionell mot antalet individer av denna art, taget med en viss koefficient beroende på arter (modeller av interspecifik konkurrens).
5. Om en art livnär sig på mat som finns tillgänglig i obegränsade mängder, är ökningen av antalet arter per tidsenhet proportionell mot antalet arter.
6. Om en art livnär sig på mat som finns tillgänglig i begränsade mängder, så regleras dess reproduktion av matkonsumtionens hastighet, d.v.s. per tidsenhet är ökningen proportionell mot mängden mat som äts.
Analogier med kemisk kinetik
Dessa hypoteser har nära paralleller med kemisk kinetik. I ekvationerna för populationsdynamik, liksom i ekvationerna för kemisk kinetik, används "kollisionsprincipen", när reaktionshastigheten är proportionell mot produkten av koncentrationerna av de reagerande komponenterna.
Faktiskt, enligt Volterras hypoteser, hastigheten bearbeta Utrotningen av varje art är proportionell mot antalet arter. Inom kemisk kinetik motsvarar detta en monomolekylär reaktion av nedbrytningen av ett visst ämne, och i en matematisk modell motsvarar det negativa linjära termer på ekvationernas högra sida.
Enligt begreppen kemisk kinetik är hastigheten för den bimolekylära reaktionen av interaktion mellan två ämnen proportionell mot sannolikheten för kollision av dessa ämnen, dvs. produkten av deras koncentration. På samma sätt, i enlighet med Volterras hypoteser, är reproduktionshastigheten för rovdjur (rovdjursdöd) proportionell mot sannolikheten för möten mellan rovdjur och bytesindivider, dvs. produkten av deras antal. I båda fallen förekommer bilinjära termer i modellsystemet på högra sidan av motsvarande ekvationer.
Slutligen motsvarar de linjära positiva termerna på högra sidan av Volterra-ekvationerna, som motsvarar tillväxten av populationer under obegränsade förhållanden, de autokatalytiska termerna för kemiska reaktioner. Denna likhet mellan ekvationer i kemiska och miljömässiga modeller tillåter oss att tillämpa samma forskningsmetoder för matematisk modellering av populationskinetik som för system av kemiska reaktioner.
Klassificering av typer av interaktioner
I enlighet med Volterras hypoteser, samspelet mellan två arter, vars antal x 1 och x 2 kan beskrivas med ekvationerna:
(9.1)
Här är parametrarna a i - konstanter för artens egen tillväxthastighet, c i‑ konstanter för självbegränsning av antal (intraspecifik konkurrens), b ij- artinteraktionskonstanter, (i, j= 1,2). Tecken på dessa koefficienter bestämmer typen av interaktion.
I den biologiska litteraturen klassificeras interaktioner vanligtvis efter de inblandade mekanismerna. Mångfalden här är enorm: olika trofiska interaktioner, kemiska interaktioner som existerar mellan bakterier och planktonalger, interaktioner av svampar med andra organismer, följd av växtorganismer förknippade särskilt med konkurrens om solljus och med utvecklingen av jordar, etc. Denna klassificering verkar enorm.
E . Odum, med hänsyn till de modeller som föreslagits av V. Volterra, föreslog en klassificering inte efter mekanismer, utan efter resultat. Enligt denna klassificering ska samband bedömas som positiva, negativa eller neutrala beroende på om förekomsten av en art ökar, minskar eller förblir oförändrad i närvaro av en annan art. Sedan kan huvudtyperna av interaktioner presenteras i tabellform.
TYPER AV INTERAKTION AV ART
|
SYMBIOS |
b 12 ,b 21 >0 |
||
|
KOMMENSALISME |
b 12 ,>0, b 21 =0 |
||
|
ROVDÖR-OFFER |
b 12 ,>0, b 21 <0 |
||
|
AMENSALISM |
b 12 ,=0, b 21 <0 |
||
|
KONKURRENS |
b 12 , b 21 <0 |
||
|
NEUTRALISM |
b 12 , b 21 =0 |
Den sista kolumnen visar tecknen på interaktionskoefficienterna från systemet (9.1)
Låt oss titta på huvudtyperna av interaktioner
TÄVLINGSEKVATIONER:
Som vi såg i föreläsning 6 är tävlingsekvationerna:
 (9.2)
(9.2)
Stationära systemlösningar:
(1).
![]()
Ursprunget för koordinater, för alla systemparametrar, är en instabil nod.
(2).
![]() (9.3)
(9.3)
C stationärt tillstånd (9.3) är en sadel vid a 1 >b 12 /Med 2 och
stabil nod vid a 1 12 /s 2 . Detta tillstånd innebär att en art dör ut om dess egen tillväxthastighet är mindre än ett visst kritiskt värde.
(3).
![]() (9.4)
(9.4)
C stationär lösning (9.4)¾ sadel kl a 2 >b 21 /c 1 och en stabil nod vid a 2< b 21 /c 1
(4).
![]() (9.5)
(9.5)
Stationärt tillstånd (9.5) kännetecknar samexistensen av två konkurrerande arter och representerar en stabil nod om förhållandet är uppfyllt:
![]()
Detta innebär ojämlikheten:
b 12
b 21
så att vi kan formulera villkoret för arters samexistens:
Produkten av koefficienterna för inter-populationsinteraktion är mindre än produkten av koefficienter inom populationsinteraktionen.
Låt faktiskt de naturliga tillväxthastigheterna för de två arterna under övervägandea 1 , a 2 är samma. Då blir det nödvändiga villkoret för stabilitet
c 2 > b 12 ,c 1 >b 21 .
Dessa ojämlikheter visar att en ökning av storleken på en konkurrent undertrycker dess egen tillväxt mer än tillväxten hos en annan konkurrent. Om antalet av båda arterna begränsas, delvis eller helt, av olika resurser, är ovanstående ojämlikheter giltiga. Om båda arterna har exakt samma behov, kommer en av dem att vara mer livskraftig och ersätta sin konkurrent.
Beteendet hos systemets fasbanor ger en tydlig uppfattning om de möjliga resultaten av konkurrensen. Låt oss likställa den högra sidan av ekvationerna i systemet (9.2) med noll:
x 1 (a 1 – c 1 x 1 – b 12 x 2) = 0 (dx 1 /dt = 0),
x 2 (a 2 –b 21 x 1 – c 2 x 2) = 0 (dx 2 /dt = 0),
I det här fallet får vi ekvationer för systemets huvudsakliga isokliner
x 2 = – b 21 x 1 / c 2 +a 2/c 2, x 2 = 0
– ekvationer för isokliner för vertikala tangenter.
x 2 = – c 1 x 1 / b 12+ a 1 /b 12 , x 1 = 0
– ekvationer för isokliner för vertikala tangenter. Punkterna för parvis skärningspunkt mellan isokliner i vertikala och horisontella tangentsystem representerar stationära lösningar av ekvationssystemet (9.2.) och deras koordinater ![]() är stationära antal konkurrerande arter.
är stationära antal konkurrerande arter.
Den möjliga placeringen av huvudisoklinerna i systemet (9.2) visas i Fig. 9.1. Ris. 9.1Amotsvarar artens överlevnadx 1, fig. 9.1 b– artens överlevnadx 2, fig. 9.1 V– samexistens av arter när villkor (9.6) är uppfyllt. Figur 9.1Gdemonstrerar triggersystemet. Här beror utfallet av konkurrensen på de ursprungliga förutsättningarna. Det stationära tillståndet som inte är noll (9.5) för båda typerna är instabilt. Detta är sadeln genom vilken separatrixen passerar och skiljer överlevnadsområdena för varje art.

Ris. 9.1.Placeringen av de viktigaste isoklinerna på fasporträttet av Volterra-konkurrenssystemet av två typer (9.2) med olika parametrar. Förklaringar i texten.
För att studera artkonkurrens genomfördes experiment på en mängd olika organismer. Vanligtvis väljs två närbesläktade arter ut och odlas tillsammans och separat under strikt kontrollerade förhållanden. Med vissa intervaller genomförs en fullständig eller selektiv folkräkning. Data från flera replikatexperiment registreras och analyseras. Studier utfördes på protozoer (särskilt ciliater), många arter av skalbaggar av släktet Tribolium, drosophila och sötvattenskräftdjur (daphnia). Många experiment har utförts på mikrobiella populationer (se föreläsning 11). Experiment utfördes också i naturen, bland annat på planar (Reynolds), två arter av myror (Pontin), etc. I fig. 9.2. skildrar tillväxtkurvorna för kiselalger som använder samma resurs (upptar samma ekologiska nisch). När den odlas i monokultur Asterionella Formosa når en konstant nivå av densitet och håller koncentrationen av resursen (silikat) på en konstant låg nivå. B. När den odlas i monokultur Synedrauina beter sig på liknande sätt och håller silikatkoncentrationen på en ännu lägre nivå. B. Under samodling (i duplikat) Synedruina förskjuter Asterionella formosa. Tydligen Synedra

Ris. 9.2.Konkurrens i kiselalger. A - när den odlas i monokultur Asterionella Formosa når en konstant nivå av densitet och håller koncentrationen av resursen (silikat) på en konstant låg nivå. b - när den odlas i monokultur Synedrauina beter sig på liknande sätt och håller silikatkoncentrationen på en ännu lägre nivå. V - med samodling (i duplikat) Synedruina förskjuter Asterionella formosa. Tydligen Synedra vinner tävlingen på grund av sin förmåga att mer fullt ut utnyttja substratet (se även föreläsning 11).
Experimenten för att studera konkurrens av G. Gause är allmänt kända, som visar överlevnaden för en av de konkurrerande arterna och gör det möjligt för honom att formulera "lagen om konkurrensutslagning". Lagen säger att endast en art kan existera i en ekologisk nisch. I fig. 9.3. Resultaten av Gauses experiment presenteras för två arter av Parametium, som upptar samma ekologiska nisch (fig. 9.3 a, b) och arter som upptar olika ekologiska nischer (fig. 9.3 c).
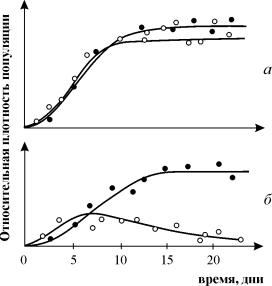
Ris. 9.3. A- Populationstillväxtkurvor för två arter Parametium i enartade grödor. Svarta cirklar – P Aurelia, vita cirklar – P. Caudatum
b- Tillväxtkurvor för P Aurelia och P . Caudatum i en blandad kultur.
Av Gause, 1934
Konkurrensmodellen (9.2) har nackdelar, i synnerhet följer det att samexistens mellan två arter är möjlig endast om deras antal begränsas av olika faktorer, men modellen anger inte hur stora skillnaderna måste vara för att säkerställa långsiktig samexistens . Samtidigt är det känt att för långvarig samexistens i en föränderlig miljö krävs en skillnad som når en viss storlek. Genom att introducera stokastiska element i modellen (till exempel att införa en resursanvändningsfunktion) kan vi kvantitativt undersöka dessa frågor.
PREDATOR+VICTIM-system
 (9.7)
(9.7)
Här, till skillnad från (9.2), tecknen b 12 Och b 21 är olika. Som i fallet med konkurrens, ursprunget
![]() (9.8)
(9.8)
är en speciell punkt av typen instabil nod. Tre andra möjliga stationära tillstånd:
![]() ,(9.9)
,(9.9)
![]() (9.10)
(9.10)
![]() (9.11)
(9.11)
Således är det möjligt för endast bytet att överleva (9,10), endast rovdjuret (9,9) (om det har andra födokällor) och samexistensen av båda arterna (9,11). Vi har redan övervägt det sista alternativet i föreläsning 5. Möjliga typer av fasporträtt för predator-bytessystemet presenteras i fig. 9.4.
Isokliner av horisontella tangenter är raka linjer
x 2 = – b 21 X 1 /c 2 + a 1/c 2, X 2 = 0,
och isokliner av vertikala tangenter- hetero
x 2 = – c 1 X 1 /b 12 + a 2 /b 12 , X 1 = 0.
Stationära punkter ligger i skärningspunkten mellan vertikala och horisontella tangentisokliner.

Från fig. 9.4 följande syns. Predator-bytes system (9.7) kan ha en stabil jämviktsposition, i vilken o Rompopulationen av offer dog helt ut ( ) och bara rovdjuren fanns kvar (punkt 2 i fig. 9.4 A). Uppenbarligen kan en sådan situation endast realiseras om, utöver den aktuella typen av offer, X 1 rovdjur X 2 – har ytterligare strömkällor. Detta faktum återspeglas i modellen av den positiva termen på höger sida av ekvationen för x2. Särskilda punkter(1) och (3) (Fig. 9.4 A) är instabila. Andra möjligheten – ett stabilt stationärt tillstånd där populationen av rovdjur helt har dött ut och bara byten finns kvar – stabil punkt(3) (Fig. 9.4 6 ). Det finns en speciell poäng här (1) – också en instabil nod.
Slutligen den tredje möjligheten – hållbar samexistens av populationer av rovdjur och bytesdjur (Fig. 9.4 V), vars stationära tal uttrycks med formlerna (9.11).
Som i fallet med en population (se föreläsning 3), för modellen (9.7) Det är möjligt att utveckla en stokastisk modell, men den kan inte lösas explicit. Därför kommer vi att begränsa oss till allmänna överväganden. Låt oss till exempel anta att jämviktspunkten ligger på ett visst avstånd från var och en av axlarna. Sedan för fasbanor på vilka värdenax 1 , x 2 förbli tillräckligt stor, kommer en deterministisk modell att vara ganska tillfredsställande. Men om någon variabel vid något tillfälle i fasbanan inte är särskilt stor, kan slumpmässiga fluktuationer bli signifikanta. De leder till det faktum att den representerande punkten flyttas till en av axlarna, vilket innebär utrotning av motsvarande art.
Således visar sig den stokastiska modellen vara instabil, eftersom den stokastiska "driften" förr eller senare leder till att en av arterna utrotas. I denna typ av modell dör rovdjuret så småningom ut, antingen av en slump eller för att dess bytespopulation först elimineras. Den stokastiska modellen av predator-bytessystemet förklarar Gauses experiment väl (Gause, 1934), i vilka ciliater Paramettum candatum tjänat som offer för ett annat ciliat Didinium nasatum – rovdjur. Förväntas enligt deterministiska ekvationer (9.7) Jämviktssiffrorna i dessa experiment var ungefär bara fem individer av varje art, så det är inte förvånande att i varje upprepat experiment dog antingen rovdjuren eller bytet (och efter dem rovdjuren) ut ganska snabbt. Resultaten av experimenten presenteras i fig. 9.5.

Ris. 9.5. Höjd Parametium caudatum och rovflikar Dadinium nasutum. Från : Gause G.F. Kampen för tillvaron. Baltimore, 1934
Så, analysen av Volterra-modeller av artinteraktion visar att, trots den stora variationen av typer av beteenden hos sådana system, kan det inte finnas odämpade fluktuationer i antal i modellen av konkurrerande arter alls. Sådana svängningar observeras emellertid i naturen och i experiment. Behovet av deras teoretiska förklaring var ett av skälen till att modellbeskrivningar formulerades i en mer generell form.
Generaliserade modeller för interaktion av två typer
Ett stort antal modeller har föreslagits för att beskriva interaktionen mellan arter, vars högra sida av ekvationerna var funktioner av antalet interagerande populationer. Frågan om att utveckla generella kriterier för att fastställa vilken typ av funktioner som kan beskriva beteendet hos den tillfälliga befolkningsstorleken, inklusive stabila fluktuationer, löstes. Den mest kända av dessa modeller tillhör Kolmogorov (1935, reviderad artikel - 1972) och Rosenzweig (1963).
 (9.12)
(9.12)
Modellen inkluderar följande antaganden:
1) Predatorer interagerar inte med varandra, d.v.s. rovdjurs reproduktionshastighet k 2 och antalet offer L utrotas per tidsenhet av ett rovdjur är inte beroende av y.
2) Ökningen av antalet bytesdjur i närvaro av rovdjur är lika med ökningen i frånvaro av rovdjur minus antalet byten som utrotats av rovdjur. Funktioner k 1 (x), k 2 (x), L(x), - är kontinuerliga och definierade på den positiva halvaxeln x, y³ 0.
3) dk 1 /dx< 0. Detta innebär att bytesreproduktionshastigheten i frånvaro av ett rovdjur minskar monotont med en ökning av antalet bytesdjur, vilket återspeglar den begränsade tillgången på mat och andra resurser.
4) dk 2 /dx> 0, k 2 (0) < 0 < k 2 (¥ ). Med en ökning av antalet offer minskar reproduktionskoefficienten för rovdjur monotont med en ökning av antalet offer, och går från negativa värden (när det inte finns något att äta) till positiva.
5) Antalet bytesdjur som förstörts av ett rovdjur per tidsenhet L(x)> 0 på N> 0; L(0)=0.
Möjliga typer av fasporträtt av systemet (9.12) presenteras i fig. 9,6:

Ris. 9.6.Fasporträtt av Kolmogorov-systemet (9.12), som beskriver interaktionen mellan två typer vid olika förhållanden av parametrar. Förklaringar i texten.
Stationära lösningar (det finns två eller tre) har följande koordinater:
(1). ` x=0;` y=0.
Ursprunget för koordinaterna för alla parametervärden är en sadel (fig. 9.6 a-d).
(2). ` x=A,` y=0.(9.13)
Abestäms utifrån ekvationen:
k 1 (A)=0.
Stationär lösning (9.13) är en sadel om B< A (Fig. 9.6 A, b, G), B bestäms utifrån ekvationen
k 2 (B)=0
Punkt (9.13) placeras i den positiva kvadranten if B>A . Detta är en stabil nod .
Det sista fallet, som motsvarar rovdjurets död och bytesdjurets överlevnad, visas i fig. 9.6 V.
(3). ` x=B,` y=C.(9.14)
Värdet på C bestäms från ekvationerna:
Punkt (9.14) – fokus (Fig. 9.6 A) eller nod (Fig.9.6 G), vars stabilitet beror på kvantitetens teckens
s 2 = – k 1 (B) – k 1 (B)B+L(B)C.
Om s>0, en punkt är stabil oms<0 ‑ точка неустойчива, и вокруг нее могут существовать предельные циклы (рис. 9.6 b)
I utländsk litteratur betraktas en liknande modell som föreslagits av Rosenzweig och MacArthur (1963) oftare:
 (9.15)
(9.15)
Var f(x) - förändringstakt i antalet offer x i frånvaro av rovdjur, F( x,y) - predationsintensitet, k- Koefficient som karakteriserar effektiviteten av att bearbeta biomassa från bytesdjur till biomassa från rovdjur. e- rovdjursdödlighet.
Modellen (9.15) reduceras till ett specialfall av Kolmogorov-modellen (9.12) under följande antaganden:
1) antalet rovdjur begränsas endast av antalet byten,
2) hastigheten med vilken ett givet rovdjur äter bytesdjur beror endast på tätheten av bytespopulationen och beror inte på tätheten hos rovdjurspopulationen.
Sedan tar ekvationerna (9.15) formen.

Vid beskrivning av samspelet mellan verkliga arter specificeras ekvationernas högra sida i enlighet med idéer om biologiska verkligheter. Låt oss överväga en av de mest populära modellerna av denna typ.
Modell av interaktion mellan två typer av insekter (MacArthur, 1971)
Modellen, som vi kommer att överväga nedan, användes för att lösa det praktiska problemet med att kontrollera skadliga insekter genom att sterilisera hanarna av en av arterna. Baserat på de biologiska egenskaperna hos artinteraktion skrevs följande modell
 (9.16)
(9.16)
Här x,y- biomassa av två typer av insekter. De trofiska interaktionerna mellan arterna som beskrivs i denna modell är mycket komplexa. Detta bestämmer formen av polynom på höger sida av ekvationerna.
Låt oss titta på den högra sidan av den första ekvationen. Insektarter Xäta artens larver på(medlem +k 3 y), men vuxna av arten påäta artens larver X föremål för hög artöverflöd X eller på eller båda typerna (medlemmar – k 4 xy, – y 2). Vid liten X arters dödlighet X högre än dess naturliga ökning (1 –k 1 +k 2 x–x 2 < 0 vid liten X). I den andra ekvationen termen k 5 speglar artens naturliga tillväxt y; –k 6 y – självbehärskning av denna typ,–k 7 x– äta larver av arten på insektsarter x, k 8 xy – ökning av arternas biomassa på på grund av konsumtion av vuxna insekter av arten på artens larver X.
I fig. 9.7 en gränscykel presenteras, vilket är banan för en stabil periodisk lösning av systemet (9.16).

Lösningen på frågan om hur man säkerställer samexistensen av en befolkning med dess biologiska miljö, kan naturligtvis inte erhållas utan att ta hänsyn till särdragen hos ett visst biologiskt system och en analys av alla dess inbördes samband. Samtidigt tillåter studiet av formella matematiska modeller oss att svara på några allmänna frågor. Det kan hävdas att för modeller som (9.12) beror faktumet om populationers kompatibilitet eller inkompatibilitet inte på deras ursprungliga storlek, utan bestäms endast av artens interaktion. Modellen hjälper till att svara på frågan: hur man kan påverka biocenosen och hantera den för att snabbt förstöra de skadliga arterna.
Förvaltningen kan reduceras till en kortsiktig, abrupt förändring av befolkningens värden X Och u. Denna metod motsvarar kontrollmetoder såsom engångsdestruktion av en eller båda populationerna med kemiska medel. Från uttalandet formulerat ovan är det tydligt att för kompatibla populationer kommer denna kontrollmetod att vara ineffektiv, eftersom systemet med tiden återigen kommer att nå en stationär regim.
Ett annat sätt är att ändra typen av interaktionsfunktioner mellan vyer, till exempel när man ändrar värdena för systemparametrar. Det är denna parametriska metod som biologiska bekämpningsmetoder motsvarar. Sålunda, när steriliserade hanar introduceras, minskar den naturliga befolkningstillväxten. Om vi samtidigt får en annan typ av fasporträtt, ett där det bara finns ett stabilt stationärt tillstånd med noll skadedjurstal, kommer kontrollen att leda till önskat resultat – förstörelse av populationen av en skadlig art. Det är intressant att notera att det ibland är tillrådligt att applicera effekten inte på skadedjuret själv, utan på sin partner. I allmänhet är det omöjligt att säga vilken metod som är mer effektiv. Detta beror på tillgängliga kontroller och på den explicita formen av funktionerna som beskriver interaktionen mellan populationer.
Modell av A.D. Bazykin
Den teoretiska analysen av modeller för artinteraktioner utfördes mest omfattande i A.D. Bazykins bok "Biophysics of Interacting Populations" (M., Nauka, 1985).
Låt oss överväga en av rovdjur-bytesmodellerna som studeras i den här boken.
 (9.17)
(9.17)
System (9.17) är en generalisering av den enklaste Volterra rovdjur-bytesmodellen (5.17) med hänsyn till effekten av rovdjursmättnad. Modell (5.17) utgår från att intensiteten av bytesbetet ökar linjärt med ökande bytetäthet, vilket inte överensstämmer med verkligheten vid höga bytetätheter. Olika funktioner kan väljas för att beskriva ett rovdjurs diets beroende av bytesdensitet. Det är viktigast att den valda fungerar med tillväxt x tenderade asymptotiskt till ett konstant värde. Modellen (9.6) använde ett logistiskt beroende. I Bazykins modell är hyperbeln vald som en sådan funktion x/(1+px). Låt oss komma ihåg att detta är formen av Monod-formeln, som beskriver beroendet av tillväxthastigheten för mikroorganismer på koncentrationen av substratet. Här fungerar bytet som substrat och rovdjuret som mikroorganismer. .
System (9.17) beror på sju parametrar. Antalet parametrar kan minskas genom att ersätta variabler:
x® (A/D)x; y ® (A/D)/y;
t® (1/A)t; g (9,18)
och beror på fyra parametrar.
För en komplett kvalitativ studie är det nödvändigt att dela upp det fyrdimensionella parameterutrymmet i områden med olika typer av dynamiskt beteende, d.v.s. bygga ett parametriskt eller strukturellt porträtt av systemet.
Sedan är det nödvändigt att konstruera fasporträtt för vart och ett av områdena i det parametriska porträttet och beskriva de bifurkationer som uppstår med fasporträtten vid gränserna för olika områden av det parametriska porträttet.
Konstruktionen av ett komplett parametriskt porträtt utförs i form av en uppsättning "skivor" (projektioner) av ett lågdimensionellt parametriskt porträtt med fasta värden för några av parametrarna.
Parametriskt porträtt av system (9.18) för fast g och liten e presenteras i fig. 9.8. Porträttet innehåller 10 områden med olika typer av beteende av fasbanor.
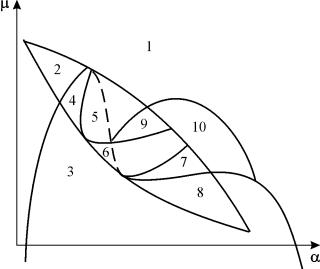
Ris. 9.8.Parametriskt porträtt av system (9.18) för fastg
och liten e
Systemets beteende vid olika förhållanden av parametrar kan vara avsevärt olika (fig. 9.9). Systemet tillåter:
1) en stabil jämvikt (regionerna 1 och 5);
2) en stabil gränscykel (regionerna 3 och 8);
3) två stabila jämvikter (region 2)
4) stabil gränscykel och instabil jämvikt inuti den (regionerna 6, 7, 9, 10)
5) stabil gränscykel och stabil jämvikt utanför den (region 4).
I de parametriska områdena 7, 9, 10 begränsas området för attraktion av jämvikt av en instabil gränscykel som ligger inuti en stabil. Den mest intressanta strukturen är fasporträttet, motsvarande område 6 i det parametriska porträttet. Det visas i detalj i fig. 9.10.

Attraktionsområdet för jämvikt B 2 (skuggat) är en "snigel" som vrider sig från det instabila fokuset B 1. Om det är känt att systemet vid det första ögonblicket var i närheten av B 1, så är det möjligt att bedöma om motsvarande bana kommer att nå jämvikten B 2 eller en stabil gränscykel som omger de tre jämviktspunkterna C (sadeln) ), B 1 och B 2 baserat på probabilistiska överväganden.

Fig.9.10.Fasporträtt av system 9.18 för parametrisk region 6. Attraktionsområde B 2 är skuggad
I ett parametriskt porträtt(9.7) det finns 22 olika bifurkationsgränser som bildas 7 olika typer av bifurkationer. Deras studie tillåter oss att identifiera möjliga typer av systembeteende när dess parametrar ändras. Till exempel när man flyttar från området 1 till område 3 födelsen av en liten gränscykel inträffar, eller den mjuka födelsen av självsvängningar runt en enda jämvikt I. En liknande mjuk födelse av självsvängningar, men kring en av jämvikterna, nämligen B 1 , uppstår när man korsar gränserna för regioner 2 och 4. När man lämnar området 4 till område 5 stabil gränscykel runt en punktB 1 "spricker" på slingan av separatriser och den enda attraktionspunkten förblir jämvikt B 2 etc.
Av särskilt intresse för praktiken är naturligtvis utvecklingen av kriterier för ett systems närhet till bifurkationsgränser. Faktum är att biologer är väl medvetna om egenskapen "buffring" eller "flexibilitet" hos naturliga ekologiska system. Dessa termer hänvisar vanligtvis till ett systems förmåga att absorbera yttre påverkan. Så länge intensiteten av den yttre påverkan inte överstiger ett visst kritiskt värde, genomgår inte systemets beteende kvalitativa förändringar. På fasplanet motsvarar detta systemets återgång till ett stabilt jämviktstillstånd eller till en stabil gränscykel, vars parametrar inte skiljer sig mycket från den ursprungliga. När intensiteten av påverkan överstiger den tillåtna nivån "bryts systemet samman" och går in i ett kvalitativt annorlunda dynamiskt beteende, till exempel dör det helt enkelt ut. Detta fenomen motsvarar en bifurkationsövergång.
Varje typ av bifurkationsövergång har sina egna särdrag, som gör det möjligt att bedöma faran för en sådan övergång för ekosystemet. Här är några allmänna kriterier som indikerar närheten till en farlig gräns. Som i fallet med en art, om, när antalet av en av arterna minskar, systemet "fastnar" nära en instabil sadelpunkt, vilket uttrycks i en mycket långsam återställning av antalet till det ursprungliga värdet, då systemet är nära den kritiska gränsen. En faroindikator är också en förändring i formen av fluktuationer i antalet rovdjur och bytesdjur. Om svängningar som är nära harmoniska blir avslappnings-svårigheter och svängningarnas amplitud ökar, kan detta leda till att systemets stabilitet försämras och att en av arterna försvinner.
Ytterligare fördjupning av den matematiska teorin om interaktion mellan arter går i linje med att detaljera strukturen hos själva populationerna och ta hänsyn till tidsmässiga och rumsliga faktorer.
Litteratur.
Kolmogorov A.N. Kvalitativ studie av matematiska modeller av populationsdynamik. // Problem med cybernetik. M., 1972, nummer 5.
MacArtur R. Grafisk analys av ekologiska system // Biologisk avdelningsrapport Perinceton University. 1971
A.D. Bazykin "Biofysik för interagerande populationer." M., Nauka, 1985.
V. Volterra: "Matematisk teori om kampen för tillvaron." M.. Vetenskap, 1976
Gause G.F. Kampen för tillvaron. Baltimore, 1934.
Populationsdynamik är en av grenarna av matematisk modellering. Det är intressant eftersom det har specifika tillämpningar inom biologi, ekologi, demografi och ekonomi. Det finns flera grundläggande modeller i det här avsnittet, varav en, "Predator-Prey"-modellen, diskuteras i den här artikeln.
Det första exemplet på en modell inom matematisk ekologi var modellen som föreslagits av V. Volterra. Det var han som först övervägde modellen för förhållandet mellan rovdjur och byte.
Låt oss överväga problemformuleringen. Låt det finnas två typer av djur, varav den ena slukar den andra (rovdjur och bytesdjur). I det här fallet görs följande antaganden: bytesresursernas matresurser är inte begränsade och därför, i frånvaro av ett rovdjur, ökar bytespopulationen enligt en exponentiell lag, medan rovdjur, separerade från sina offer, gradvis dör av hunger, också enligt en exponentiell lag. När rovdjur och bytesdjur börjar leva i närheten av varandra, blir förändringar i deras populationsstorlekar relaterade till varandra. I detta fall kommer uppenbarligen den relativa ökningen av antalet bytesdjur att bero på storleken på rovdjurspopulationen och vice versa.
I denna modell antas det att alla rovdjur (och alla byten) är i samma förhållanden. Samtidigt är offrens matresurser obegränsade, och rovdjur livnär sig uteslutande på offren. Båda populationerna lever i ett begränsat område och interagerar inte med några andra populationer, och det finns inga andra faktorer som kan påverka populationens storlek.
Själva den matematiska "rovdjur-byte"-modellen består av ett par differentialekvationer som beskriver dynamiken hos populationer av rovdjur och bytesdjur i sitt enklaste fall, när det finns en population av rovdjur och en av rovdjur. Mönstret kännetecknas av fluktuationer i storleken på båda populationerna, med toppen hos rovdjur något bakom toppen hos bytesdjur. Denna modell finns i många arbeten om populationsdynamik eller matematisk modellering. Det har täckts brett och analyserats med matematiska metoder. Men formler kanske inte alltid ger en uppenbar uppfattning om processen som äger rum.
Det är intressant att ta reda på exakt hur populationsdynamiken i denna modell beror på de initiala parametrarna och hur mycket detta motsvarar verkligheten och sunt förnuft, och att se detta grafiskt utan att tillgripa komplexa beräkningar. För detta ändamål, baserat på Volterra-modellen, skapades ett program i Mathcad14-miljön.
Låt oss först kontrollera modellen för överensstämmelse med verkliga förhållanden. För att göra detta, låt oss överväga degenererade fall när endast en av populationerna lever under givna förhållanden. Det har visats teoretiskt att i frånvaro av rovdjur ökar bytespopulationen oändligt med tiden, och rovdjurspopulationen i frånvaro av bytet dör ut, vilket generellt motsvarar modellen och den verkliga situationen (med den angivna formuleringen av problem).
De erhållna resultaten återspeglar de teoretiska: rovdjur dör gradvis ut (fig. 1), och antalet byten ökar oändligt (fig. 2).
Fig. 1 Beroende av antalet rovdjur i tid i frånvaro av bytesdjur

Fig. 2 Beroende av antalet bytesdjur i tid i frånvaro av rovdjur
Som framgår motsvarar systemet i dessa fall den matematiska modellen.
Låt oss överväga hur systemet beter sig under olika initiala parametrar. Låt det finnas två populationer - lejon och antiloper - rovdjur respektive bytesdjur, och initiala indikatorer ges. Då får vi följande resultat (Fig. 3):
Tabell 1. Systemoscillerande modskoefficienter

Fig.3 System med parametervärden från Tabell 1
Låt oss analysera erhållna data baserat på graferna. Med den initiala ökningen av antiloppopulationen observeras en ökning av antalet rovdjur. Observera att toppökningen i populationen av rovdjur observeras senare, under nedgången i populationen av rovdjur, vilket är ganska förenligt med verkliga begrepp och den matematiska modellen. Faktum är att en ökning av antalet antiloper innebär en ökning av matresurserna för lejon, vilket innebär en ökning av deras antal. Vidare leder den aktiva konsumtionen av antiloper av lejon till en snabb minskning av antalet bytesdjur, vilket inte är förvånande, med tanke på rovdjurets aptit, eller snarare frekvensen av rovdjur som äter bytesdjur. En gradvis minskning av antalet rovdjur leder till en situation där bytespopulationen befinner sig i gynnsamma förhållanden för tillväxt. Sedan upprepas situationen med en viss period. Vi drar slutsatsen att dessa förhållanden inte är lämpliga för individers harmoniska utveckling, eftersom de medför kraftiga minskningar av bytespopulationen och kraftiga ökningar i båda populationerna.
Låt oss nu ställa in predatorns initiala antal lika med 200 individer samtidigt som andra parametrar bibehålls (fig. 4).
Tabell 2. Systemoscillerande modskoefficienter

Fig.4 System med parametervärden från Tabell 2
Nu svänger systemet mer naturligt. Under dessa antaganden existerar systemet ganska harmoniskt, det finns inga kraftiga ökningar och minskningar av antalet nummer i båda populationerna. Vi drar slutsatsen att med dessa parametrar utvecklas båda populationerna tillräckligt jämnt för att leva tillsammans i samma territorium.
Låt oss ställa in det initiala antalet rovdjur till 100 individer, antalet bytesdjur till 200, samtidigt som andra parametrar bibehålls (fig. 5).
Tabell 3. Systemoscillerande modskoefficienter

Fig.5 System med parametervärden från Tabell 3
I det här fallet är situationen nära den första situationen. Observera att med en ömsesidig ökning av populationerna har övergångarna från en ökning till en minskning av bytespopulationen blivit mjukare, och rovdjurspopulationen förblir i frånvaro av bytesdjur med ett högre numeriskt värde. Vi drar slutsatsen att när en population är nära besläktad med en annan, sker deras interaktion mer harmoniskt om de specifika initiala populationerna är tillräckligt stora.
Låt oss överväga att ändra andra systemparametrar. Låt de initiala siffrorna motsvara det andra fallet. Låt oss öka reproduktionshastigheten för offer (Fig. 6).
Tabell 4. Systemoscillerande modskoefficienter

Fig.6 System med parametervärden från Tabell 4
Låt oss jämföra detta resultat med resultatet som erhölls i det andra fallet. I detta fall observeras en snabbare tillväxt av offret. I det här fallet beter sig både rovdjuret och bytet som i det första fallet, vilket förklarades av den låga populationsstorleken. Med denna interaktion toppar båda populationerna vid värden som är mycket högre än i det andra fallet.
Låt oss nu öka tillväxthastigheten för rovdjur (Fig. 7).
Tabell 5. Systemoscillerande modskoefficienter

Fig.7 System med parametervärden från Tabell 5
Låt oss jämföra resultaten på samma sätt. I detta fall förblir systemets allmänna egenskaper desamma, med undantag för förändringen i period. Som förväntat blev perioden kortare, vilket förklaras av den snabba minskningen av predatorpopulationen i frånvaro av bytesdjur.
Och slutligen, låt oss ändra koefficienten för interspecifik interaktion. Låt oss först öka frekvensen av rovdjur som äter bytesdjur:
Tabell 6. Systemoscillerande modskoefficienter

Fig.8 System med parametervärden från Tabell 6
Eftersom rovdjuret äter sitt byte oftare har den maximala populationsstorleken ökat jämfört med det andra fallet, och skillnaden mellan den maximala och minsta populationsstorleken har också minskat. Systemets svängningsperiod förblir densamma.
Och låt oss nu minska frekvensen av rovdjur som äter bytesdjur:
Tabell 7. Systemoscillerande modskoefficienter

Fig.9 System med parametervärden från Tabell 7
Nu äter rovdjuret bytet mer sällan, den maximala populationsstorleken har minskat jämfört med det andra fallet, och den maximala populationsstorleken för bytet har ökat med 10 gånger. Av detta följer att bytespopulationen under dessa förhållanden har större frihet när det gäller reproduktion, eftersom rovdjuret behöver mindre massa för att få nog av den. Skillnaden mellan högsta och lägsta befolkningsstorlek har också minskat.
När man försöker modellera komplexa processer i naturen eller samhället, på ett eller annat sätt, uppstår frågan om modellens riktighet. Naturligtvis, vid modellering, förenklas processen och några mindre detaljer försummas. Å andra sidan finns det en fara att förenkla modellen för mycket och därigenom slänga ut viktiga egenskaper hos fenomenet tillsammans med de oviktiga. För att undvika denna situation, innan modellering är det nödvändigt att studera ämnesområdet där denna modell används, undersöka alla dess egenskaper och parametrar, och viktigast av allt, lyfta fram de funktioner som är de mest betydande. Processen ska ha en naturlig beskrivning, intuitivt förståelig, som i huvudsak sammanfaller med den teoretiska modellen.
Den modell som beaktas i detta arbete har ett antal betydande nackdelar. Till exempel antagandet om obegränsade resurser för offret, frånvaron av tredjepartsfaktorer som påverkar dödligheten för båda arterna, etc. Alla dessa antaganden återspeglar inte den verkliga situationen. Men trots alla brister har modellen fått stor spridning på många områden, även långt ifrån ekologi. Detta kan förklaras av det faktum att "rovdjur-byte"-systemet ger en allmän uppfattning om samspelet mellan arter. Interaktioner med miljön och andra faktorer kan beskrivas med andra modeller och analyseras tillsammans.
Relationer av typen "rovdjur-byte" är ett väsentligt inslag i olika typer av livsaktiviteter där det sker en kollision mellan två interagerande parter. Denna modell äger rum inte bara inom ekologi, utan även inom ekonomi, politik och andra verksamhetsområden. Till exempel är ett av områdena relaterade till ekonomi analysen av arbetsmarknaden, med hänsyn till tillgängliga potentiella arbetstagare och lediga jobb. Detta ämne skulle vara en intressant fortsättning på arbetet med rovdjur-bytesmodellen.
Låta och vara antalet bytesdjur respektive rovdjur. Låt oss anta att den enda begränsande faktorn som begränsar reproduktionen av bytesdjur är trycket på dem från rovdjur, och reproduktionen av rovdjur begränsas av mängden mat de får (antalet bytesdjur). Då, i frånvaro av rovdjur, bör antalet bytesdjur växa exponentiellt med en relativ hastighet a, och rovdjur i frånvaro av bytesdjur bör också dö ut exponentiellt med en relativ hastighet t. Koefficienterna a och m är koefficienterna för naturlig ökning bytesdjur respektive naturlig dödlighet hos rovdjur.
Låt vara antalet (eller biomassa) bytesdjur som konsumeras av ett rovdjur per tidsenhet, och en del av energin som erhålls från denna biomassa spenderas av rovdjuret på reproduktion, och resten spenderas på att upprätthålla basal metabolism och jaktaktivitet. Då kan ekvationen för predator-bytessystemet skrivas som

Funktionen brukar kallas rovdjurets trofiska funktion eller rovdjurets funktionella svar (funktionell respons) på bytesdjurets befolkningstäthet. Det är dessa funktioner som vanligtvis identifieras i experimentella studier av predation, och det är nu accepterat att dessa funktioner vanligtvis tillhör någon av följande tre typer (Fig. 11). Uppenbarligen beror systemets dynamiska beteende till stor del på typen av trofisk funktion.

Ris. 11. Olika typer av trofiska funktioner i rovdjur-bytessystemet: a) denna typ är karakteristisk för ryggradslösa djur och vissa arter av rovfiskar; b) en trofisk funktion med en uttalad mättnadströskel är karakteristisk för filtermatande rovdjur (till exempel många blötdjur); c) denna typ är karakteristisk för ryggradsdjur - organismer som uppvisar ganska komplext beteende (till exempel kapabla att lära). Den trofiska funktionen kommer att ha ett liknande utseende om byten kan utveckla en defensiv strategi (till exempel gömma sig i ett skydd otillgängligt för rovdjur).
För små värden på x, till exempel, när trofiska relationer i systemet är spända och nästan alla byten blir byte för ett rovdjur som alltid är hungrig och inte blir mätt (situationen är ganska vanlig i naturen), den trofiska funktion kan betraktas som en linjär funktion av antalet byten, d.v.s. Dessutom, låt oss anta att . Sedan

System (2.2) sammanfaller, upp till notation, med den klassiska predator-bytesmodellen av V. Volterra, som visade att detta system har en integral av formen
![]()
där If är de initiala värdena för antalet bytesdjur respektive rovdjur, då
![]()
och ekvation (2.3) beskriver en familj av slutna kurvor kapslade i varandra, motsvarande fasbanorna för periodiska lösningar av systemet (2.2).

Ris. 12. Fasporträtt av det klassiska Volterra-systemets rovdjur - byte.
Observera att när C ökar, ökar amplituderna för oscillationerna och y (fig. 12). Vid minimivärdet dras dessa kurvor samman till en punkt med koordinater. Det är lätt att se att de är lösningar på system (2.2) för , d.v.s. dess icke-triviala jämvikt. För fallet med små oscillationer nära detta tillstånd kan modellekvationerna skrivas som
![]()
Rötterna till motsvarande karakteristiska ekvation är, dvs punkten är mitten. En period av små svängningar, där svängningarna i antalet av en art förskjuts i fas i förhållande till svängningarna hos en annan av
I system (2.2) finns ytterligare en jämviktsposition - ursprunget. Det är lätt att se att denna punkt är en sadel. Koordinataxlarna är separatriser, där axeln går in i sadeln och axeln lämnar den (se fig. 12).
Trots att Volterra-modellen kunde förklara många faktiskt observerade fenomen, har den en stor nackdel - ojämnheten (i ordets matematiska betydelse) i Volterra-cyklerna, så att för alla, hur svaga, störningar som helst av faskoordinaterna går systemet från en cykel till en annan. Tydligen borde mer adekvata modeller ha denna "grovhet" egenskap.
Från stabilitetsteorins synvinkel är systemets jämviktstillstånd (x, y) ett tillstånd av likgiltig jämvikt, stabilt enligt Lyapunov, men inte asymptotiskt. Frånvaron av asymptotisk jämviktsstabilitet indikerar det i Volterra-systemet det finns inga mekanismer som försöker bevara dess icke-triviala jämviktstillstånd.
Ovanstående uttalande om jämviktsstabiliteten bevisas ganska lätt genom att konstruera motsvarande Lyapunov-funktion. Men mer om det i nästa stycke.

Liknande artiklar



