Späť v 20. rokoch. A. Lotka a o niečo neskôr nezávisle od neho V. Volter navrhli matematické modely, ktoré popisujú konjugované kolísanie počtu predátorov a koristi.
Model pozostáva z dvoch komponentov:
C – počet predátorov; N – počet obetí;
Predpokladajme, že bez prítomnosti predátorov bude populácia koristi rásť exponenciálne: dN/dt = rN. Korisť však predátori ničia rýchlosťou, ktorá je určená frekvenciou stretnutí medzi predátorom a korisťou a frekvencia stretnutí sa zvyšuje so zvyšujúcim sa počtom predátorov (C) a koristi (N). Presný počet nájdenej a úspešne zjedenej koristi bude závisieť od účinnosti, s akou dravec korisť nájde a uloví, t.j. z a’ – „účinnosť vyhľadávania“ alebo „frekvencia útokov“. Frekvencia „úspešných“ stretnutí medzi predátorom a korisťou, a teda aj miera jedenia obetí, sa teda bude rovnať a’СN a vo všeobecnosti: dN/dt = rN – a’CN (1*).
Pri nedostatku potravy jednotliví predátori chudnú, hladujú a umierajú. Predpokladajme, že v uvažovanom modeli sa veľkosť populácie predátora pri nedostatku potravy v dôsledku hladovania exponenciálne zníži: dC/dt = - qC, kde q je úmrtnosť. Smrť je kompenzovaná narodením nových jedincov rýchlosťou, o ktorej sa v tomto modeli verí, že závisí od dvoch okolností:
1) miera spotreby potravín, a’CN;
2) účinnosť (f), s ktorou táto potrava prechádza do potomstva dravca.
Plodnosť predátora sa teda rovná fa’CN a vo všeobecnosti: dC/dt = fa’CN – qC (2*). Rovnice 1* a 2* tvoria Lotka-Wolterov model. Vlastnosti tohto modelu je možné študovať, skonštruovať líniové izokliny, ktoré zodpovedajú konštantnej veľkosti populácie, a pomocou takýchto izoklinál možno určiť správanie interagujúcich populácií dravec-korisť.
V prípade populácie koristi: dN/dt = 0, rN = a’CN alebo C = r/a’. Pretože r a a’ = const, izoklina pre obeť bude priamka, pre ktorú je hodnota C konštantná:
Pri nízkej hustote predátora (C) počet koristi (N) stúpa, naopak klesá.
Podobne pre dravce (rovnica 2*) s dC/dt = 0, fa’CN = qC, alebo N = q/fa’, t.j. Izoklinála pre predátora bude čiara, pozdĺž ktorej je N konštantné: Pri vysokej hustote koristi sa veľkosť populácie predátora zvyšuje a pri nízkej hustote klesá.
Ich počet podlieha neobmedzeným konjugovaným fluktuáciám. Pri veľkom počte koristi sa zvyšuje počet predátorov, čo vedie k zvýšeniu tlaku predátorov na populáciu koristi a tým k zníženiu jej počtu. Tento pokles následne vedie k obmedzeniu dravcov v potrave a poklesu ich počtu, čo spôsobuje oslabenie tlaku predátorov a zvýšenie počtu koristi, čo opäť vedie k zvýšeniu populácie predátorov. , atď.
Populácie vykonávajú rovnaký cyklus kmitov neobmedzene dlho, kým nejaký vonkajší vplyv nezmení ich počet, po čom populácie vykonávajú nové cykly neobmedzených kmitov. V skutočnosti sa životné prostredie neustále mení a populácie sa budú neustále posúvať na novú úroveň. Aby boli cykly oscilácií, ktoré populácia robí, pravidelné, musia byť stabilné: ak vonkajší vplyv zmení úroveň populácie, potom musia smerovať k pôvodnému cyklu. Takéto cykly sa nazývajú stabilné, limitné cykly.
Model Lotka-Wolter nám umožňuje ukázať hlavný trend vo vzťahu dravec – korisť, ktorý sa prejavuje vo výskyte výkyvov v populácii koristi sprevádzaných výkyvmi v populácii dravca. Hlavným mechanizmom takýchto výkyvov je časový posun, ktorý je vlastný postupnosti stavov od vysokého počtu koristi k vysokému počtu predátorov, potom k nízkemu počtu koristi a nízkemu počtu predátorov, k vysokému počtu koristi, atď.
5) POPULAČNÉ STRATÉGIE PREDÁTOROV A PRIMÁTOV
Vzťah „predátor – korisť“ predstavuje prepojenia v procese prenosu hmoty a energie z fytofágov do zoofágov alebo od predátorov nižšieho rádu k predátorom vyššieho rádu. Autor: Na základe povahy týchto vzťahov sa rozlišujú tri typy predátorov:
A) zberači. Predátor zbiera malé, pomerne početné mobilné obete. Tento typ predácie je typický pre mnohé druhy vtákov (kulík, plutvy, jazvičky a pod.), ktoré vynakladajú energiu len na hľadanie obetí;
b) skutočných predátorov. Dravec prenasleduje a zabíja korisť;
V) pastierov. Tieto predátory opakovane využívajú korisť, napríklad gadfly alebo konské muchy.
Stratégia získavania potravy medzi predátormi je zameraná na zabezpečenie energetickej účinnosti výživy: energetický výdaj na získavanie potravy by mal byť menší ako energia získaná pri jej asimilácii.
Skutoční predátori sa delia na
„ženci“, ktorí sa živia bohatými zdrojmi (vrátane planktónových rýb a dokonca aj veľrýb), a „lovci“, ktorí hľadajú menej hojnú potravu. Vo svojom poradí
„Poľovníci“ sa delia na „útočníkov“, ktorí číhajú na korisť (napríklad šťuka, jastrab, mačka, mantis chrobák), „hľadači“ (hmyzožravé vtáky) a „prenasledovateľov“. Pre druhú skupinu si hľadanie potravy nevyžaduje veľké výdavky na energiu, ale na zmocnenie sa koristi (levy v savanách) jej treba veľa. Niektoré dravce však dokážu kombinovať prvky stratégie rôznych možností lovu.
Rovnako ako vo vzťahu „fytofág-rastlina“ nie je v prírode pozorovaná situácia, v ktorej sú všetky obete zjedené predátormi, čo v konečnom dôsledku vedie k ich smrti. Ekologická rovnováha medzi predátormi a korisťou je udržiavaná špeciálnymi mechanizmami, čím sa znižuje riziko úplného vyhladenia obetí. Takže obete môžu:
Utečte pred predátorom. V tomto prípade sa v dôsledku adaptácií zvyšuje mobilita obetí aj predátorov, čo je typické najmä pre stepné zvieratá, ktoré sa pred prenasledovateľmi nemajú kam skryť;
Získajte ochrannú farbu („predstierajte“ listy alebo vetvičky) alebo naopak jasnú farbu, N.: červená, upozorňujúca dravca na horkú chuť. Je dobre známe, že farba zajaca sa mení v rôznych obdobiach roka, čo mu umožňuje maskovať sa v tráve v lete a na pozadí bieleho snehu v zime. Adaptívne zmeny farby sa môžu vyskytnúť v rôznych štádiách ontogenézy: mláďatá tuleňov sú biele (farba snehu) a dospelí jedinci sú čierni (farba skalnatého pobrežia);
Rozdeľte do skupín, čím je ich hľadanie a chytanie pre dravca energeticky náročnejšie;
Skryť sa v prístreškoch;
Prejdite na aktívne obranné opatrenia (bylinožravce s rohmi, ostnaté ryby), niekedy spoločné (pižmoň môže zaujať „všestrannú obranu“ pred vlkmi atď.).
Predátori si zase rozvíjajú nielen schopnosť rýchlo prenasledovať korisť, ale aj čuch, ktorý im umožňuje určiť polohu koristi podľa čuchu. Mnoho druhov predátorov roztrháva nory svojich obetí (líšky, vlci).
Sami zároveň robia všetko pre to, aby sa vyhli odhaleniu ich prítomnosti. To vysvetľuje čistotu malých mačiek, ktoré trávia veľa času toaletou a zahrabávaním exkrementov, aby eliminovali zápach. Dravce nosia „maskovacie rúcha“ (pruhovanie šťúk a ostriežov, vďaka čomu sú menej nápadné v húštinách makrofytov, pruhoch tigrov atď.).
Nedochádza ani k úplnej ochrane všetkých jedincov v populáciách dravých zvierat pred predátormi, pretože by to viedlo nielen k úhynu hladujúcich predátorov, ale v konečnom dôsledku ku katastrofe populácií koristi. Zároveň sa pri absencii alebo znížení populačnej hustoty predátorov zhoršuje genofond populácie koristi (choré a staré zvieratá zostávajú zachované) a v dôsledku prudkého nárastu ich počtu je podkopaná ponuka potravy.
Z tohto dôvodu sa zriedkavo pozoruje vplyv závislosti veľkosti populácie koristi a predátorov - pulzácia veľkosti populácie koristi, po ktorej nasleduje pulzácia veľkosti populácie predátora s určitým oneskorením („Lotka-Volterra efekt“). .
Medzi biomasou predátorov a korisťou je stanovený pomerne stabilný pomer. R. Ricklefs teda poskytuje údaje, že pomer biomasy predátora a koristi sa pohybuje od 1:150 do 1:300. V rôznych ekosystémoch mierneho pásma Spojených štátov amerických pripadá na jedného vlka 300 malých belorítok (hmotnosť 60 kg), 100 veľkých jeleňov wapiti (hmotnosť 300 kg) alebo 30 losov (hmotnosť 350). Rovnaký vzor bol nájdený v savanách.
Pri intenzívnom využívaní fytofágnych populácií ľudia z ekosystémov často vylučujú predátorov (napr. vo Veľkej Británii srnčia a jelenia zver, ale nie vlci, v umelých nádržiach, kde sa chovajú kapry a iné rybničné ryby, nie sú šťuky). V tomto prípade úlohu predátora vykonáva samotná osoba, ktorá odstraňuje časť jedincov populácie fytofágov.
Špeciálny variant predácie sa pozoruje u rastlín a húb. V rastlinnej ríši existuje asi 500 druhov, ktoré sú schopné chytať hmyz a čiastočne ho tráviť pomocou proteolytických enzýmov. Dravé huby tvoria lapacie zariadenia vo forme malých oválnych alebo guľovitých hláv umiestnených na krátkych vetvách mycélia. Najbežnejším typom pasce sú však adhézne trojrozmerné siete pozostávajúce z veľkého počtu krúžkov vytvorených v dôsledku vetvenia hýf. Dravé huby dokážu chytiť pomerne veľké zvieratá, ako sú škrkavky. Potom, čo sa červ zamotá do hýf, rastú vo vnútri tela zvieraťa a rýchlo ho naplnia.
1. Konštantná a priaznivá úroveň teploty a vlhkosti.
2. Hojnosť potravy.
3.Ochrana pred nepriaznivými faktormi.
4. Agresívne chemické zloženie biotopu (tráviace šťavy).
1. Prítomnosť dvoch biotopov: prostredie prvého rádu je hostiteľský organizmus, prostredie druhého rádu je vonkajšie prostredie.
Izv. univerzity "PND", ročník 19, č. 2, 2011
FENOMÉN MATEMATICKÉHO MODELU LOTKA-VOLTERRA A JEMU PODOBNÉ*
DI. Trubetskov
Matematický model Lotka-Volterra (často nazývaný model „predátor-korisť“) je použiteľný na opis rôznych procesov v biológii, ekológii, medicíne, sociálnom výskume, histórii, rádiofyzike a iných vedách.
Tento prehľad, ktorý má do značnej miery metodologický charakter, skúma varianty tohto modelu a podobných modelov vo vzťahu k analýze množstva prírodných a spoločenských javov.
Diskutuje sa o nasledujúcich modeloch: modely interakcie medzi znečistením a životným prostredím; model triedneho boja; model beztriednej spoločnosti z éry lovcov a zberačov; model vojenských operácií; vírusový model infekčnej choroby; model šírenia epidémií vrátane modelu infekcie počítačovým vírusom; model interakcie kognitívnych a/alebo emocionálnych režimov mozgu.
Kľúčové slová: matematický model Lotka-Volterra, systém dravec-korisť, analýza prírodných a spoločenských javov.
"...Pri riešení právnych prípadov, ako pri stanovovaní diagnózy lekárom alebo pri pokusoch vedených chemikom, zohráva rozhodujúcu úlohu rozpoznanie typických vzorov."
Bernhard Schlink. Návrat**
Úvod na environmentálnych príkladoch
Model Lotka-Volterra je vo svojej podstate matematickým popisom darwinovského princípu boja o existenciu, ktorý Charles Darwin vo svojej autobiografii uvádza nasledovne. „Po preštudovaní mnohých okolností som dospel k záveru, že existuje boj o existenciu. Raz mi napadlo, že za podobných okolností sa priaznivé druhy budú ďalej rozvíjať
"Petrohrad: Vydavateľská skupina "ABC-Classics", 2010, s. 67
Cesta, ktorou sa Darwin uberal, je v knihe dobre opísaná. Je zvláštne, že Darwin bol konečne presvedčený o svojich záveroch po prečítaní knihy „Essay on the Law of Population“ od Thomasa Roberta Malthusa. Táto esej zohrala dôležitú úlohu v spoločenskom a politickom živote Británie v 90. rokoch 19. storočia, keďže bola vnímaná ako rozumné pochopenie prirodzených zákonov spoločnosti. V 30. rokoch 20. storočia malthusiánske doktríny potláčali vládnu politiku. Pripomeňme si Malthusove argumenty, ktoré sú prekvapivo jednoduché. Keďže podľa Malthusa má ľudstvo prirodzenú tendenciu zväčšovať svoj druh, produkcia potravín nemohla zostať na rovnakej úrovni.
„Aj keď existuje približná rovnováha, ako tvrdí autor: množstvo ľudí zomiera v dôsledku prírodných faktorov (hlad alebo choroby) alebo ľudských činov (vojna, sexuálna abstinencia alebo hriešne činy, ako je zabíjanie novorodencov), je to nevyhnutné z dôvodu celkovej rovnováhy existencie,
Malthus išiel ďalej a tvrdil: ... tento údel (smrť) je údelom najslabšieho, najchudobnejšieho a najchorľavejšieho člena spoločnosti. Nie je možné uniknúť pripravenému osudu, teda vôli Božej. Jeden jednoduchý záver naznačoval sám, ale autor na to varoval „Ak dáte almužnu chudobným, ľudská rasa sa rozrastie a bude nedostatok jedla,“
Zdalo sa, že viktoriánske časy potvrdili, že Malthus mal pravdu. Vyskytli sa potravinové nepokoje, diskusie o chudobných zákonoch a verejný odpor proti obilným zákonom. V roku 1834 viktoriánsky establishment v reakcii na zmeny chudobného zákona zriadil chudobince, kde ľudia pracovali za chlieb a vodu ako alternatívu k cirkevným sociálnym spoločnostiam.
Darwin žil v tomto svete, bol jeho predstaviteľom a pohyboval sa v rovnakých kruhoch ako Malthus. Malthusove myšlienky rozširuje na všetko živé: v prírode prebieha boj o život, boj o existenciu, v ktorom najskôr zomiera najhorší a najslabší organizmus a víťazia rozvinutejšie formy, zdravšie a prispôsobenejšie. Práve títo jedinci rodia potomkov. Organizmy sa rýchlejšie prispôsobujú podmienkam prostredia, ak sa podobné prípady boja v určitých intervaloch opakujú.
Začnime našu ďalšiu prezentáciu (na pripomenutie) modelmi interagujúcich populácií – prvými matematickými modelmi Lotky a Volterry. Budeme sa riadiť prezentáciou kapitoly 5 knihy, pretože je vhodná pre ďalší popis modelov interakcie medzi znečistením a životným prostredím.
Autori poukazujú predovšetkým na prácu Volterra, pričom poznamenávajú, že podobný model „navrhla Lotka o niečo skôr (Volterra vykonal oveľa kompletnejšiu analýzu tohto systému, Lotka sa zakaždým snažila zdôrazniť svoju prioritu pri zaznamenávaní tohto modelu) “,
Model Lotka-Volterra popisuje interakciu dvoch druhov – populácie predátorov a populácie koristi.
Nech N(t) je počet koristi, P(t) je počet predátorov v čase t. Potom má model Lotka-Volterra podobu:
AN - bNP, - = -dP + cNP, (1)
a, b, c, d sú kladné konštanty.
Pripomeňme, že sústava rovníc (1) je založená na nasledujúcich predpokladoch:
V neprítomnosti predátorov sa korisť neobmedzene rozmnožuje podľa rovnice dN/dt = aN, ktorá sa niekedy nazýva Malthusova rovnica;
Predátori bez obetí vymierajú podľa rovnice dP/dt = -dP;
Pojmy úmerné súčinu NP sa považujú za premenu energie jedného zdroja na energiu druhého (vplyv vplyvu populácie dravcov na populáciu koristi, teda výsledkom ich stretnutia, je znížiť rýchlosť rastu dN/dt počtu koristi o množstvo NP, úmerné počtu predátorov).
Ak zavediete nové premenné
() N(t)d () P(t)b
n(x) = -v(x) = -^,
potom systém rovníc (1) nadobudne tvar
N - nv, - = YV(n - 1), dx dx
Sústava rovníc (2) má dva pevné body: (0, 0) - sedlo a (1,1),
pre ktoré sú vlastné hodnoty Jacobiánskej matice A
- ^1,2 = ±iVУ,
i = l/- 1, to znamená, že rovnovážna poloha (1.1) je nehyperbolická a na základe lineárnej analýzy nie je možné vyvodiť záver o jej stabilite.
Fázové trajektórie systému rovníc (2), ako je známe, sú integrálnymi krivkami rovnice
dv (n - 1)v dn Y (1 - v)n'
ktorého riešenie
yn + v - ln nYv = H.
Dá sa ukázať (pozri napr.), že v blízkosti (1, 1) úrovňových čiar funkcie H sú uzavreté krivky zodpovedajúce periodickým riešeniam sústavy rovníc (2) (obr. 1). Konštanta H vo výraze (3) je určená počiatočnými podmienkami, napríklad n(0) = n0 a v(0) = vo.
Matematický model opísaný sústavou rovníc (2) je štruktúrny
je nestabilný, to znamená, že malá zmena na pravej strane v ňom môže viesť ku kvalitatívnej zmene v správaní riešenia. Toto je jeho hlavná nevýhoda. Navyše, preto je ťažké hovoriť o zhode teórie a experimentu.
Jedným z dôvodov štrukturálnej nestability modelu je, že je konzervatívny, to znamená, že má prvý integrál. Ako zdôrazňuje autor knihy, napriek naznačenej hlavnej nevýhode modelu systém rovníc (1) umožňuje vyvodiť netriviálne závery potvrdené mnohými pozorovaniami. Predovšetkým je formulovaný Volterrov princíp: ak v systéme „predátor-korisť“ opísanom modelom (1) sú oba druhy vyhubené rovnomerne a v pomere k počtu ich jedincov, potom sa priemerný počet obetí zvyšuje a priemerný počet predátorov klesá.
Kniha poskytuje dva príklady existencie tohto účinku v prírode. Jedným z nich je sociálny fakt, ktorý viedol Volterra k jeho teórii boja o existenciu: počas prvej svetovej vojny bol rybolov v Jadranskom mori značne obmedzený, čo na prekvapenie biológov viedlo k nárastu predátorov a poklesu počet koristi. Druhý príklad je menej známy. Podľa Volterrovho princípu sa ukazuje dvojaký charakter použitia repelentov proti hmyzu (insekticídov) na konzervovanie plodín na poliach. Takmer všetky takéto chemikálie pôsobia nielen na škodcov, ale aj na ich prirodzených nepriateľov, čo často vedie k zvýšeniu počtu škodcov a zníženiu napríklad vtákov, ktoré sa týmito škodcami živia. Podľa Volterrovho princípu je teda v ekosystéme dravec-korisť populácia koristi citlivejšia na proces proporcionálneho znižovania jedincov v populácii.
V terminológii V.I. Arnolda, model opísaný systémom rovníc (1) sa nazýva „rigidný“. Mimochodom, časť knihy V.I. Arnold „Tvrdé“ a „mäkké“ matematické modely (M.: MCNO, 2000) sa nazývajú „Tvrdé modely ako cesta k chybným predpovediam“. S malou zmenou v modeli zmäkne, keď sa na pravú stranu (1) pridajú malé výrazy, berúc do úvahy napríklad súťaž medzi korisťou o potravu alebo predátormi o korisť (uvedené sú špecifické typy rovníc nižšie). Potom vo všeobecnom tvare možno rovnicu (1) prepísať takto:
V závislosti od malých korekcií ef (P, N) a napr. (P, N) sa napríklad scenáre znázornené na obr. 2.
Z hľadiska výkladu je zaujímavý Obr. 2, b. Ak sú P banditi a N robotníci, vývoj systému podľa Arnolda vedie buď k prudkému nárastu počtu banditov, alebo k ich takmer úplnému zániku, keďže robotníkov okradli natoľko, že už nezostane nič. zobrat. Systém nakoniec spadá do oblasti hodnôt P a N tak veľkých alebo malých, že model sa stáva nepoužiteľným, pretože sa menia zákony evolúcie.
Tu je niekoľko komplikovanejších modelov predátor-korisť.
Systém „predátor-korisť“ zohľadňujúci vnútrodruhovú konkurenciu.
aN - bPN + ef (P, N)
dP + cPN + napr.(P,N), napr< 1.
Ryža. 2. a - rovnovážny stav je stabilný: za akýchkoľvek počiatočných podmienok sa po dlhom čase nastolí rovnaký stav; b - rovnovážny stav B je nestabilný: systém „predáva“; c - v systéme s nestabilným stacionárnym stavom A sa v priebehu času nastaví periodický režim C (na rozdiel od systému (1) (pozri obr. 1) prebiehajú kmity určitej amplitúdy, nezávisle od maličkosti výchylky)
kde K\,2 sú potenciálne kapacity ekologických systémov, ktoré sú určené dostupným množstvom zdrojov a zodpovedajú hraničným hodnotám počtu obyvateľov. V bezrozmerných premenných má tento systém rovníc tvar
kde N(t) = (a/c)u(x), P(t) = av(x)/b, t Model konkurencie
at, y = d/a, a = a/(cK\), V = d/(aK2).
it1=1-Ш-eNN w=1-f)-hNN (5)
Tu sú n, r2, e, h kladné konštanty.
Analýza sústavy rovníc (5) ukazuje: ak n populácií lineárne závisí od m zdrojov a m< n, то по крайней мере одна из популяций вымирает.
Model mutualizmu (symbióza)
ui(ri + aiiui + ai2u2),
u2 (Г2 + a2iui + a22u2).
Na opísanie vzájomnosti stačí to vyžadovať
ai2 > 0, a2i > 0.
Podrobný rozbor vyššie uvedených modelov nájdete napríklad v knihách a.
Na konci časti, po prezentácii, uvažujeme zovšeobecnený model Lotka-Volterra, popísaný systémom rovníc
A(u) - B(u,v), - = -D(v) + C (u,v), (7)
kde u av sú počty koristi a predátorov, A(u) je reprodukčná funkcia koristi v neprítomnosti predátorov; D(v) - funkcia vyhynutia predátorov v neprítomnosti koristi; funkcia B(u, v) opisuje požieranie koristi predátormi; C(u, v) je účinnosť spotreby koristi predátormi. Uveďme možné ďalšie faktory vnútro- a medzipopulačných vzťahov.
Nelineárna závislosť miery reprodukcie populácií koristi od hustoty pri nízkych hodnotách hustoty (nedostatok dostatočného počtu páriacich sa párov): A(u) = au2/(N + u), kde a a N sú kladné konštanty.
Vnútrodruhová konkurencia koristi: A(u) = au(l - u/K).
Sýtosť predátorov: Bi(u) = bu/(1 + au) - trofická funkcia predátora.
Nelineárny charakter predácie predátormi: Bi(u) = bu2/(1 + au).
Súťaž medzi predátormi o korisť: B2(v) = bv/(1 + |3v). V tomto prípade B(u,v) = Bi(u)B2(v).
Súťaž predátorov o zdroje odlišné od koristi: C(v) = v/(1 + v/Ki).
Nelineárny charakter závislosti miery reprodukcie dravca od hustoty populácie pri nízkych hodnotách hustoty: C2(v) = cv/(Nv + v), pričom C(u,v) = C1(u)C2(v) a C1(u) = B1(u), D2(v) = cv, to znamená C(u,v) = = cv/(Nv + v) Bi(u).
1. Matematický model interakcie znečistenia s prostredím, interpretovaný ako model „predátor-korisť“
Situáciu „znečistenie – príroda“ možno interpretovať ako špeciálny prípad modelu „predátor – korisť“, keď príroda vystupuje ako obeť a znečistenie ako predátor. Hlavným predpokladom modelu je, že životné prostredie aktívne absorbuje a spracováva znečistenie do určitej hranice.
Z kvalitatívnych úvah sú v systéme životného prostredia a znečistenia možné nasledujúce tri zásadne odlišné scenáre interakcie.
1. Pri malých emisiách znečistenia ho životné prostredie úplne recykluje (trvalo udržateľná situácia).
2. S nárastom emisií znečistenia, v závislosti od vonkajších podmienok a náhodných príčin, môže byť životné prostredie v uspokojivom stave, alebo môže zaniknúť (bistabilná situácia)*.
3. Napokon tretia situácia zodpovedá environmentálnej katastrofe – úplnému vyhynutiu prírody.
Predpokladajme, že všeobecné pozadie znečistenia a stav životného prostredia možno charakterizovať nasledujúcimi premennými: koncentrácia znečistenia P a hustota biomasy E, resp. Ak existuje trvalý zdroj znečistenia, proces vývoja znečistenia možno opísať rovnicou
kde a je výkon zdroja znečistenia za jednotku času, b je koeficient prirodzeného zničenia znečistenia (takéto zničenie sa nazýva „mŕtve“, resp.
„Bistabilná situácia môže nastať, keď v blízkosti niektorých podnikov zostáva vegetačný kryt len miestami. Vyplýva to z pozorovaní a v mnohých smeroch môže slúžiť ako oneskorený signál na prijatie urgentných opatrení na záchranu prírody.
prirodzený rozptyl). Za počiatočnej podmienky P(t = 0) = Po má riešenie rovnice (8) tvar:
P (t) = b+(Po - D e~bt'
to znamená, že v priebehu času koncentrácia znečistenia prirodzene klesá.
Ďalej predpokladajme, že znečistenie je v neustálej interakcii s prostredím, čo má na znečistenie čistiaci účinok. Za uzavretý budeme považovať aj systém „životné prostredie – znečistenie“. Potom možno proces interakcie s prostredím opísať nasledujúcim systémom rovníc:
J- = a - bP- f (E, P), - = g(E) - h(E, P), (9)
kde funkcia f (E, P) > 0 - opisuje absorpciu a spracovanie znečistenia životného prostredia; g(E) je termín popisujúci dynamiku prostredia bez znečistenia; h(E, P) je funkcia, ktorá popisuje škodlivé účinky znečistenia na životné prostredie.
Zoberme si f (E, P) = cEP a h(E, P) = dEP ako funkcie interakcie medzi znečistením a voľne žijúcimi zvieratami, kde c a d sú konštantné koeficienty.
Budeme predpokladať, že pri absencii znečistenia možno správanie prostredia opísať logistickou rovnicou, teda g(E) = rE(1 - E/K), kde r je konštantný koeficient a K zodpovedá do maximálnej hodnoty E pri dE/dt = 0 Dosadením výrazov pre f (E, P), h(E, P) a g(E) do sústavy rovníc (9) dostaneme
^ = a - bP-cEP, = rE (1-E) - dEP.
Prechod v sústave rovníc (10) k bezrozmerným premenným
T = bt, a = -pr, uo
získame najjednoduchší matematický model interakcie znečistenia s prostredím vo forme nasledujúcich rovníc:
A - u - uv, dt
v(u0 - u) - pv2.
Je ľahké vidieť, že systém rovníc (10) je systémom predátor-korisť, kde korisť je znečistenie (nie biologický objekt) a biologicky aktívne prostredie je predátor.
V systéme rovníc (11) možno parameter a interpretovať ako zovšeobecnený výkon zdroja znečistenia; uo je maximálna povolená koncentrácia pre daný ekosystém (ak u > u0, dv/dt< 0, и природа вымирает); p - характеристика экосистемы - коэффициент внутривидовой конкуренции природы.
Rovnovážne polohy sústavy rovníc (11), zodpovedajúce podmienkam du/dt = 0 a dv/dt = 0, sa dajú ľahko nájsť z rovníc.
a - u(l + v) = 0, -v(u - uo) - pv2 = 0.
Potom Ai = (a, 0); A2 ((uo + p + Q)/2, (uo - p - Q)/(2p));
A3 ((uo + p - Q)/2, (uo - p + Q)/(2p)), Q = ^/(uo + p)2 - 4ap.
Druhá a tretia rovnovážna poloha existujú, ak [(u + p)2-4aP]>0. Štandardná lineárna analýza stability indikovaných rovnovážnych polôh vedie k parametrickému portrétu sústavy rovníc (11), znázornenej na obr. 3. Na obr. Obrázok 4 ukazuje zodpovedajúce fázové portréty systému rovníc (11).
Ak parametre patria do oblasti IV, potom všetky trajektórie s výnimkou stabilnej variety Ai majú tendenciu k A3 ako t ^ x (obr. 4, a). Oblasť II zodpovedá fázovému portrétu na obr. 4, b. V oblasti III je jediný globálny atraktor A1 (obr. 4, c).
Opísaný matematický model teda zodpovedá trom situáciám kvalitatívne opísaným vyššie.
Na začiatku tejto časti však bolo uvedené ako hlavný predpoklad, že model obsahuje prahovú hodnotu znečistenia, ktorú príroda dokáže spracovať. Tento efekt je potrebné vziať do úvahy v matematickom modeli.
Ryža. 3. Parametrický portrét sústavy rovníc (11). Pre pevné u0 je celá oblasť prípustných parametrov rozdelená na štyri sady
Na tento účel kniha navrhuje použiť jednu z trofických funkcií, ktoré popisujú proces nasýtenia, napr.
Potom pri nízkej koncentrácii znečistenia f (E, P) ~ cPE, pri vysokej koncentrácii f (E, P) cE, ktorá je realistickejšia ako pôvodný výraz.
Po zredukovaní sústavy rovníc (9) do bezrozmernej podoby, berúc do úvahy vzťah (12) a preznačenia v, bola získaná nasledujúca sústava rovníc:
A - u ---, - = v(uo - u) - pv , dt k + u dt
kde k = A(d/b) > 0 popisuje mieru vplyvu prírody na znečistenie: čím väčšia je jej hodnota, tým menšia je miera absorpcie živej prírody a naopak.
V knihe sa tiež uvádza, že v sústave rovníc (13) je možný jeden, dva alebo tri rovnovážne stavy rovnakého charakteru ako v sústave (11). Okrem toho autori zdôrazňujú, že apriórne predpoklady týkajúce sa výberu určitých funkčných závislostí v tomto modeli sú adekvátnejšie realite ako vybrané modely (11).
2. Matematický model čistenia odpadových vôd
Znečisťujúcu látku budeme považovať za „korisť“ a biologicky aktívny kal za „predátora“. Proces biochemickej oxidácie znečisťujúcej látky bude interpretovaný ako jej „požieranie“ mikroorganizmami aktivovaného kalu.
Predpokladajme, že existuje stály zdroj znečistenia a aktivovaný kal je schopný spracovať znečistenie do určitej hranice. Domnievame sa, že zmena koncentrácie aktivovaného kalu v čistej vode klesá podľa exponenciálneho zákona.
Potom možno dynamiku čistenia odpadových vôd opísať rovnicami
dd _ = a - bD(P) - cf (P, E), - = -dS + eh(P, E),
kde P (t) je koncentrácia znečistenia vody, E (t) je hustota biomasy aktivovaného kalu, D(P) je funkcia rozptylu charakterizujúca prirodzený rozpad znečistenia; f (P, E) a h(P, E) - trofické funkcie charakterizujúce proces čistenia znečisťujúcich látok biologicky čistým kalom; a > 0 - výkon zdroja znečistenia; d > 0 je konštanta charakterizujúca rýchlosť poklesu aktivovaného kalu v čistej vode; c a e sú kladné konštanty.
Predpokladajme, že D(P) = P a trofické funkcie „predátora“ a „koristi“ sú rovnaké a majú tvar:
f (P, E) = h(P, E) = , kde r > 0 je konštanta.
Potom budeme mať:
A - bP--------, -
Po zavedení bezrozmerných premenných m = at, P = ru(x), E = (rd/c) v(x) má sústava rovníc (14) tvar
kde a = a/(rd), b = b/d, y = e/d. Pevné body sústavy (15) Ai(ui,vi) =
= (a/fr 0) a A2(u2, v2) = (1/(Y-1), Hy-1)-p]y/(y-1)).
Štandardná lineárna analýza ukazuje, že bod Ai(ui, vi) je sedlový bod v< a(Y - 1) и y >0 a stabilný uzol pre β > a(Y - 1).
Ak zvýšite hodnotu parametra a pri pevných hodnotách v a (Y - 1), čo zodpovedá zvýšeniu intenzity zdroja znečistenia, vznikne stabilný rovnovážny stav A^(u2,v2). Ak je parameter a malý (intenzita znečistenia je nízka), potom sa hmotnosť biologicky aktívneho kalu blíži k nule. Keďže znečisťujúca látka je potravou pre biologicky aktívny kal, pri nedostatočnej intenzite škodliviny biologicky aktívny kal zaniká.
Je rozumné predpokladať, že existuje určitá prahová hodnota koncentrácie znečistenia P *, nad ktorou klesá čistiaca schopnosť kalu, a zvážiť trofické funkcie druhu
f (P, E) = EPe-rP, r = konštanta > 0.
V tomto prípade je prahová hodnota znečistenia P* = 1/r. Reálne je v popisovanom procese ešte jedna zložka - proces prevzdušňovania - nasýtenia kalu kyslíkom, čo výrazne zvyšuje schopnosť biologicky aktívneho kalu spracovať škodlivinu. Berúc do úvahy vyššie uvedené, matematický model čistenia bude mať podobu:
a - bP - cf (P, E),
dE + ef (P, E)Q(t),
kde a, b, c,d,e = const > 0. Tu Q(t) je koncentrácia kyslíka a R> 0 určuje množstvo prítoku kyslíka do systému za jednotku času.
Analýza sústavy rovníc (16) vykonaná autormi knihy za predpokladu, že f (P, E) = Pg(E), kde g(E) je hladká monotónne rastúca nezáporná funkcia (a g(0) = 0), ukazuje, že keď s dostatočne blízkymi hodnotami parametra R, ktorý určuje množstvo kyslíka vstupujúceho do systému, je vždy možné dosiahnuť stabilitu systému.
3. Modelovanie triedneho boja
Napíšme rovnice modelu „predátor-korisť“ vo forme nasledujúceho systému:
kde x, y sú počty obyvateľov, x1 a y1 sú ich stacionárne hodnoty a aab sú konštanty.
Tento model sa používa na opis malých mestských oblastí. V tomto prípade premenná x v sústave rovníc (17) znamená plochu využitia pôdy, y - pozemková renta, a, b, xi, yi - niektoré parametre.
Zaujímavú aplikáciu modelu na opis triedneho boja uvádza Goodwin. Pozrime sa na dva typy občanov: robotníkov a kapitalistov.
Robotníci míňajú všetky svoje príjmy wL na spotrebu, kapitalisti akumulujú svoj dôchodok Y - wL, kde Y je výrobný výstup. Cena spotrebného tovaru sa normalizuje na jednu.
Nech K znamená kapitál, a = ао egt = Y/L - produktivita práce, rastúca konštantným tempom d, k = K/Y - koeficient kapitálovej náročnosti výroby, N = Noent - ponuka na trhu práce, ktorá rastie s tempom rastu n Podiel nákladov práce vo vzťahu k národnému dôchodku je wL/Y = w/a. Preto je podiel na zisku kapitalistov (1 - w/a). Keďže úsporu možno definovať ako S = Y - wL = (1 - w/a)Y, investičný podiel je dK/dt = S = (1 - w/a)Y alebo (dK/dt)(1/K) = (1-w/a)(Y/K); V tomto prípade zanedbávame odchod kapitálu do dôchodku.
Pri konštantnej hodnote kapitálovej náročnosti k = K/Y dostaneme dk/dt = 0 a
dY dL 1 dt Y dt L
Okrem toho z (18), berúc do úvahy skutočnosť, že dK/dt = (1 - w/a)L, vyplýva
dY 1 dK 1 / w \ 1
dt Y dt K V a) k-
Zo vzťahov (19) a (20) nakoniec zistíme:
Zavedieme nové premenné: y = w/a - podiel nákladov práce; x = L/N - pomer obsadenosti.
Po jednoduchých transformáciách, berúc do úvahy (21) dostaneme
dx - = x dt 1 - y k (d + p)
dt =у w dt a dt
Wf (x), lim f (x) dt
limf(x)< 0,
Lineárna aproximácia f (x) v tvare (dw/dt)(l/w) = -r + bx zmení (23) na nasledujúcu rovnicu:
R + px - g. (24)
Kombinácia (22) a Eq.
(24) sa dostávame k modelu Goodwin, opísanému systémom
y[-(g + r) + px).
Je zrejmé, že sústava rovníc (25) je ekvivalentná sústave rovníc (17), preto všetky všeobecné závery platné pre sústavu (17) platia aj pre Goodwinov model.
Všimnite si, že Goodwinov model, ktorý zohľadňuje interakciu medzi úrovňou zamestnanosti a zákonom stanoveným podielom odvodov na mzdách, veľmi pripomína klasické modely politickej ekonómie (niekedy sa mu hovorí neomarxistický). Model opäť upozorňuje na diela klasických ekonómov ako Ricardo, Smith, Marx. Bohužiaľ, model je konštrukčne nestabilný.
Model „predátor“ – „korisť“ teda možno použiť na modelovanie javov vzťahu medzi mestskou pozemkovou rentou a intenzitou využívania pôdy, nezamestnanosťou a dynamikou ekonomického rastu.
4. Model „predátor-korisť“ na opis beztriednej spoločnosti – spoločnosť z éry lovcov a zberačov
Toto obdobie je charakterizované nízkou hustotou obyvateľstva a komunálnym (kmeňovým) usporiadaním ľudského spoločenstva. Človek bol úplne závislý od prírody, prebiehal neustály boj o prežitie a hlavným vyťaženým zdrojom bola potrava. Človek bol súčasťou ekosystému a od ostatných školských predátorov sa líšil v podstate iba schopnosťou používať oheň a primitívne nástroje pre svoje potreby. Zmena veľkosti populácie v konkrétnej oblasti bola určená najmä stavom zdrojovej základne a možno ju opísať modelom „predátor-korisť“ – systémom rovníc (1). Teraz v týchto rovniciach N je veľkosť populácie lovených objektov („obete“); P je počet starých lovcov („predátorov“) v posudzovanej oblasti; cNP - miera narodenia „predátorov“; dP - miera úmrtnosti „predátorov“; aN je miera úmrtnosti „obetí“ (predpokladá sa, že ich zásoba potravín je neobmedzená); bNP je miera úhynu „koristi“ v dôsledku ich vyhubenia „predátormi“ (predpokladá sa, že toto je jediná príčina úmrtnosti „koristi“. Riešenia týchto rovníc boli analyzované v Úvode). Keďže lovci-zberači konzumovali to, čo sa v prírode vyprodukovalo bez ich účasti, veľkosť populácie bola určená vonkajšími podmienkami, ktoré nemohli ovplyvniť (v modeli počet lovcov kolíše okolo priemernej hodnoty
Po = a/b. Všimnite si, že to vysvetľuje relatívnu stabilitu populácie Zeme počas doby kamennej až do obdobia neolitu.
Keď človek domestikoval zvieratá a naučil sa pestovať plodiny, premenil sa z lovca-zberača na pastiera a farmára. Výrazne sa tak zvýšil koeficient a a rovnovážny počet ľudí, keďže hodnota koeficientu b, ktorý charakterizuje spotrebu, zostala približne na rovnakej úrovni.
5. Podobná ideológia ľudí na opis vojenských akcií
Frederick Lanchester – anglický polyhistor a inžinier – v knihe navrhol jednoduchý model boja dvoch protivníkov (povedzme dvoch armád), ktorý nesie jeho meno. Všimnite si, že rovnaký model navrhol dôstojník cárskej armády M.O. Osipov v roku 1915. Preto je spravodlivé nazývať tento model modelom Lanchester-Osipov. Ďalej v prezentácii budeme prácu sledovať. V tomto modeli je stav systému opísaný bodom (x, y) kladného kvadrantu roviny. Súradnice x a y tohto bodu predstavujú počet nepriateľských armád. Modelové rovnice sú nasledovné:
Tým, - = -ax. (26)
Tu a a b predstavujú silu zbraní armády x a armády y. Inými slovami, každý vojak armády x zabije vojaka armády y za jednotku času (a teda každý vojak armády y zabije b vojakov armády x). Celková účinnosť všetkých výstrelov armády x je teda daná sekerou a pre armádu y hodnotou. Práca predpokladá, že kontinuálna aproximácia je dostatočne dobrá a rýchlosť zmeny počtu vojakov je úmerná účinnosti nepriateľských striel. „Tvrdé“, podľa V.I. Arnold, model (26) pripúšťa presné riešenie v tvare:
ax2 - bу2 = konšt. (27)
K zmene počtu armád x a y dochádza pozdĺž hyperboly špecifikovanej rovnicou (27) (obr. 5). Akú hyperbolu bude vojna nasledovať, závisí od východiskového bodu. Hyperboly sú oddelené priamkou l/ax = \[bу. Ak začiatočný bod leží nad touto čiarou, potom hyperbola smeruje na os y. To znamená, že počas vojny sa veľkosť armády x v konečnom čase zníži na nulu. Armáda víťazí, nepriateľ je zničený. Ak začiatočný bod leží pod čiarou, vyhráva armáda x. V priamej línii, ako píše Arnold, vojna „k spokojnosti všetkých“ končí vyhladením oboch armád. To však trvá nekonečne dlho: konflikt naďalej tlčie, keď sú už obaja protivníci vyčerpaní. Zo vzťahu (27) vyplýva, že na priamke ^fax = -\[bу. Potom má prvá rovnica sústavy (26) tvar: dx/dt =
Y/abx. Jeho riešením je x = Ce~^a>t, kde
С = a je zrejmé, že x > 0 pri t > Ж Obr. 5. „Tvrdý“ model vojny
Ryža. 6. „Mäkký“ model vojny
Z modelu (26) vyplýva, že na boj s dvakrát tak početným nepriateľom je potrebné mať štyrikrát silnejšie zbrane, s trojnásobným - deväťkrát silnejším atď. Naznačujú to odmocniny v rovnici priamky.
Je jasné, že výsledný „kanibalistický“ model v Arnoldovej terminológii je značne idealizovaný a je nebezpečné ho aplikovať na reálnu situáciu. Čo sa zmení, ak
Tento systém rovníc už nie je možné riešiť explicitne. Ale model sa stáva „mäkkým“ v dôsledku zmien v a(x, y) a b(x, y). Krivky v rovine (x, y) už nie sú hyperbolami a nie sú oddelené priamkou (obr. 6). Ale hlavný kvalitatívny záver zostáva rovnaký: situácie „x vyhráva“ a „y vyhráva“ sú oddelené neutrálnou čiarou „obe armády sa navzájom zničia v nekonečnom čase“.
6. Najjednoduchší vírusový model infekčného ochorenia
V roku 1974 G. Bell navrhol najjednoduchší model imunitnej odpovede, v ktorom je interakcia medzi antigénom a protilátkou opísaná v termínoch „predátor-korisť“.
Uveďme verziu teórie G. Bella, založenú na všeobecnejšom modeli G.I. Marčuk.
Hlavné faktory v modeli infekčnej choroby sú tieto:
Koncentrácia patogénnych množiacich sa antigénov V (t);
koncentrácia protilátky F(t); Protilátky znamenajú substráty imunitného systému.
systémy, ktoré neutralizujú antigény (imunoglobulíny, bunkové receptory);
Predpokladá sa, že koncentrácia plazmatických buniek C - nosičov a producentov protilátok - je konštantná;
Neberie sa do úvahy stupeň poškodenia cieľového orgánu.
Rovnica popisujúca zmenu počtu antigénov (cudzích buniek prenikajúcich do tela) má tvar
W = eV - YFV (28)
Prvý člen v rovnici (28) je rýchlosť rastu antigénov v dôsledku reprodukcie; c - koeficient násobenia antigénu; termín yFV opisuje počet antigénov neutralizovaných F protilátkami za jednotku času; y je koeficient spojený s pravdepodobnosťou neutralizácie antigénu protilátkami pri stretnutí s nimi.
Aby sme získali druhú rovnicu, vypočítajme bilanciu počtu protilátok reagujúcich s antigénom. Bude mať
dF = pCdt - nyFVdt - c/Fdt. (29)
Prvý člen na pravej strane rovnice (29) opisuje tvorbu protilátok plazmatickými bunkami počas času dt; p je rýchlosť produkcie protilátky plazmatickou bunkou. Druhý výraz (nyFVdt) popisuje pokles počtu protilátok v dôsledku komunikácie s antigénmi a existuje n protilátok na neutralizáciu jedného antigénu. Tretí výraz (ifFdt) opisuje pokles populácie protilátok v dôsledku starnutia; if je koeficient nepriamo úmerný dobe rozpadu protilátok. Vydelením rovnice (29) dt dostaneme
RS - (ak + nyV)F. (tridsať)
Ak na t< to V(t) = 0, то начальные условия имеют вид
V (do) = Vo, F (do) = Fo. (31)
Nech je to, čo nasleduje, = 0.
Uvažujme o dvoch extrémnych prípadoch dynamiky ochorenia. Predpokladajme, že telo neprodukuje protilátky tejto špecifickosti, teda F(t) = Fo = 0 pre všetky t > 0 a p = 0. Potom z (28) vyplýva, že
- = |3V a V (t) = Voeet, (32)
kde Vo je dávka infekcie (počiatočná koncentrácia antigénov) pri t = 0. Toto riešenie samozrejme zodpovedá priebehu ochorenia s fatálnym koncom, pretože neexistujú žiadne faktory kompenzujúce rast antigénov. Druhý obmedzujúci prípad: protilátky prítomné v tele (špecifické pre daný antigén) sú dostatočné na zničenie všetkých antigénov, ktoré sa dostali do tela, bez spustenia mechanizmu tvorby protilátok. V tomto prípade v rovnici (28) v ^ yF. Za predpokladu, že dávka infekcie Vo je malá, hodnotu F možno považovať za konštantnú, určenú normálnou hladinou protilátok F*. Potom má rovnica (28) tvar
F = (v - YF*> V
a jeho riešenie je napísané takto:
V = Voe-(YF*-e)t.
V obmedzujúcom prípade, keď B = 0
V = Voe-YF h, (33)
čo zodpovedá vysokej imunologickej bariére.
Riešenia (32) a (33) sú uvedené na obr. 7.
Pre dané hodnoty modelových koeficientov a počiatočných podmienok je prirodzené, že celá rodina rôznych dynamik chorôb sa zmestí do tieňovanej oblasti na obr. 7.
limitované riešeniami pre dva limitujúce prípady
7. Podobné vzorce šírenia epidémie
Zvážte Kermack-McKendrick SIR model za predpokladu, že jednotlivci v populácii môžu byť v troch rôznych stavoch:
S (t) - zdraví jedinci, ktorí sú ohrození a môžu dostať infekciu;
I(t) - nosiči infikovaných chorôb;
R(t) - tí, ktorí sa uzdravili a prestali šíriť chorobu (do tejto kategórie patria napr. tí, ktorí získali imunitu alebo aj tí, ktorí zomreli).
Typický vývoj jednotlivca v populácii je opísaný nasledujúcim diagramom:
Zostavme fenomenologický model zodpovedajúci diagramu s použitím nasledujúcich predpokladov:
1) populácia je uzavretá a má konštantnú veľkosť N;
2) doba, počas ktorej je chorá osoba nebezpečná pre ostatných, je rozdelená podľa exponenciálneho zákona s priemerom 1/r;
3) kontakty sú náhodné a rovnako možné (rovnomerné miešanie).
Za týchto predpokladov budeme mať:
kde X je takzvaná sila infekcie, ktorá sa v tomto prípade rovná súčinu počtu kontaktov za jednotku času uskutočnených jedným zdravým človekom (toto číslo označujeme ako cN), pravdepodobnosť prenosu infekcie na ho pri kontakte p a pravdepodobnosť stretnutia s chorou osobou I(t) /N. Označme = cp. Potom dostaneme X = |3! a dostávame sa k štandardnému tvaru rovníc:
wdt = S-■<’" ~dt = -о* d + d. + dt = 0и
S(t) + I (t) + R(t) = N.
Tretia rovnica v redukovanom systéme je nadbytočná a keďže dS/dt< 0, то естественно считать фазовым пространством треугольник S(t) + I(t) < N.
Z prvej a druhej rovnice sústavy (34) vyplýva, že
dL = (M-T^ = -1+ o o = Y I = o dS eSI S' o in, I =
Ak vezmeme do úvahy počiatočné podmienky S(0) = So, I(0) = I0, nájdeme prvý integrál I + S - o ln S = I0 + S0 - o ln S0, pomocou ktorého môžeme zostrojiť fázu krivky na rovine (S, I) (obr. 8). Krivky znázornené na obr. 8, b, sa dobre zhodujú s údajmi o morovej epidémii v Bombaji v roku 1906.
Kniha navrhuje použiť model diskutovaný vyššie s určitými úpravami na analýzu infekcií počítačovými vírusmi. Vyhlásenie o probléme je nasledovné.
Nech S je počet počítačov, ktoré sú infikované vírusom, I je časť počítačov, ktoré sú infikované vírusom a nemajú antivírusový softvér, a R je časť počítačov, ktoré majú správnu antivírusovú ochranu ( imunita). Potom majú modelové rovnice tvar (pozri sústavu rovníc (34)):
^ = -aSI, ^ = aSI - bI, ^ = bI. dt dt dt
Hlavnou myšlienkou úpravy tohto klasického epidemického modelu je zavedenie prvku cyklickosti, teda prvku regulácie pridávania nových počítačov náchylných na epidémiu. V každom novom časovom období sa počet infikovaných počítačov vypočíta na základe počtu infikovaných počítačov zostávajúcich z predchádzajúceho okamihu a počet počítačov vystavených epidémii sa hrá ako náhodná veličina podľa Poissonovho zákona s rozdelením pm = (Xm/m\)e-x, m = 1,2, ...
Riešenie problému nie je dané. Autori navrhujú nasledovné riešenie. Zostrojte graf funkcie I(t) numerickým riešením vyššie uvedenej sústavy rovníc s počiatočnými podmienkami 5(0) = 100, I(0) = 1, R(0) = 0, X = 10, a = 0,23 b = 0,5. Navrhuje sa pridať nové prvky pri tk = k, k = 1, 2, ...100.
Možno niektorého z čitateľov bude zaujímať riešenie tohto problému a jeho množstvo možností.
8. Zovšeobecnený model Lotka-Volterra na popis
interakcie medzi kognitívnymi a/alebo emocionálnymi režimami mozgu
Podľa práce by sa rozsiahle kognitívne vzorce (režimy alebo reprezentácie pozorované v experimente) v prevádzkovom režime mozgu mali navzájom potláčať, čo by sa prirodzene malo vyskytovať postupne v priebehu času. Inými slovami, pracujúci mozog demonštruje kognitívnu a emocionálnu aktivitu vo forme reťazca kombinácií funkčných režimov, ktoré sa v priebehu času navzájom nahrádzajú, pričom tieto kombinácie samotné sú určené typom mentálnej aktivity.
Podľa autorov „... súťaž o konečné zdroje je jednou z hlavných operačných síl, ktoré riadia mozgové procesy. Týmito zdrojmi sú energia (kyslík a glukóza) a informácie (pozornosť a pamäť). Je dôležité, aby informačné zdroje boli produktom činnosti samotného mozgu, to znamená, že môžu ovplyvňovať emócie a kognitívne funkcie rôznymi spôsobmi (napríklad prostredníctvom riadenia pozornosti) a mali by byť zahrnuté do všeobecného dynamického modelu, ktorý definuje pracovnom priestore. ...Režimy reprezentujúce emócie a kognitívne funkcie v pracovnom priestore si navzájom vymieňajú informácie a súperia o energiu a informačné zdroje.“
funkčne určené interakciou rôznych mozgových subsystémov, ich súťaživosťou a synchronizáciou (koordináciou) v čase.“ Vyššie uvedené procesy vzájomného súperenia kognitívnych a emocionálnych režimov, ako aj emocionálnych a kognitívnych režimov navzájom, navrhujeme v práci popísať sústavami rovníc typu Lotka-Volterra v nasledujúcej zovšeobecnenej podobe :
Vi(E) -^2 Wi,j(E)xi(t)
Xi(t) tu charakterizuje aktivitu i-tého módu (veľkosť i-tej populácie v ekológii); n je počet interagujúcich režimov (populácií); u(E) - informácie vstupujúce do systému alebo dostupné zdroje; f^-(E) - prvky interakčnej matice; n(t) je multiplikatívny šum prítomný v systéme; t je charakteristický čas, ktorý určuje proces.
Ak vezmeme do úvahy špecifiká zdrojov, o ktoré súperia kognitívne a emocionálne režimy, mali by existovať tri podsystémy rovníc (36): emocionálne režimy, kognitívne režimy a zdroje (pozornosť, pamäť, energia). Úloha pozornosti je obzvlášť dôležitá. V článku sa pre zjednodušenie predpokladá ... že popis súťaženia o pozornosť medzi emóciami a kognitívnymi funkciami si nevyžaduje špecifikáciu jednotlivých režimov, a preto sa môžeme obmedziť na uvažovanie o súťaži o pozornosť „vo všeobecnosti “ emócií B = ^П = 1 Bi a „vo všeobecnosti » kognitívnych módov A = ^П=1 Ai. Potom môžu byť základné rovnice napísané ako:
Oi(I,B,D)Ra -^2 Pij(D)Aj(t)
Zi(S, A,D)Rb -£ lij (D)Bj(t)
Ai(t)n(t), i = 1,...N,
Bi(t)n(t), i = 1, ...M,
RA(t) A - (RA(t) + Fl(1, D)Rb(t)) = Rb(t) B - (Rb(t) + Fb(S, D)RA(t))
V týchto rovniciach sú Ai a Bi nezáporné premenné zodpovedajúce kognitívnym a emocionálnym režimom, ktorých celková intenzita je označená A a B; I - informácie z vonkajšieho sveta; S - charakteristika emocionálneho vplyvu z vonkajšieho sveta (v prípade negatívnych emócií - stresu); D - úroveň užívaných liekov; ta a tv sú charakteristické obdobia vedomej a emocionálnej aktivity; n(t) - multiplikačný šum; koeficienty Fa a Fv určujú intenzitu boja o pozornosť zo strany emócií a myslenia. Závislosť parametrov Oi a Zi od A a B popisuje priamy vplyv aktivity kognitívnych procesov na emócie a emócií na myslenie. V článku
počítačová analýza vyššie uvedených rovníc je prezentovaná v prítomnosti vonkajšieho stimulu S(t), ktorý charakterizuje úroveň stresu. Diskutuje sa aj o interakcii medzi náladou a myslením. Analyzuje sa interakcia troch zložiek mozgovej aktivity: správania, emocionálnej pamäte a kognitívnej aktivity pri absencii vonkajších vplyvov.
Namiesto záveru
V nádhernej knihe Yu.I. Neymark má také slová.
„Čo je to modelka? Slovo „model“ je z talianskeho jazyka a znamená „kópia, vzorka, prototyp“, jeho štúdiom spoznávame originál. To znamená, že matematický model je prototypom nejakej časti sveta – objektu, systému, zariadenia, stroja, procesu, zariadenia, ktorého štúdiom chápeme túto časť sveta...“
Ak je model štandardný (pozri epigraf) alebo, ako sa hovorí, štandardný, potom s jeho pomocou môžete pochopiť nie jednu, ale rôzne časti sveta. Presne takýmto modelom je matematický model Lotka-Volterra. Predložená recenzia samozrejme pokrýva iba časť možných aplikácií a netvrdí, že je univerzálna.
Bibliografia
1. Ch. Darvin. Autobiografia. 1958. S. 120.
2. Hnedá Janet. Charles Darwin. Pôvod druhov / Ser. "10 kníh, ktoré zmenili svet." M.: AST: as, 2009. 220 s.
3. Malthus T.R. Test na populačnom princípe, pretože ovplyvňuje budúce zlepšenie spoločnosti. 1798.
http://www.faculty.rsu.edu/felwell/Theorists/Malthus/essay2.htm
4. Lotka A. Prvky fyzikálnej biológie. Baltimore, 1925. Pretlačené Doverom v roku 1956 ako Elements of Mathematical Biology.
5. Volterra V. Matematická teória boja o existenciu / Prel. z francúzštiny M.: Nauka, Hlavná redakcia fyzikálnej a matematickej literatúry, 1976. 288 s.
6. Bratus A.S., Novožilov A.S., Platonov A.P. Dynamické systémy a modely biológie. M.: FIZMATLIT, 2010. 400 s.
7. Bazykin A.D. Nelineárna dynamika interagujúcich populácií. Moskva-Iževsk: Inštitút počítačového výskumu, 2003. 368 s.
8. Arnold V.I. „Tvrdé“ a „mäkké“ modely // Príroda. 1998. Číslo 4. P. 3.
9. Bratus A.S., Meshcherin A.S., Novozhilov A.S. Matematické modely interakcie medzi znečistením a životným prostredím // Bulletin Moskovskej štátnej univerzity. Ser. Výpočtová matematika a kybernetika. 2001. T. 6. S. 140.
10. Zang W.-B. Synergická ekonomika. Čas a zmeny v nelineárnej ekonomickej teórii / Transl. z angličtiny M.: Mir, 1999. 335 s.
11. Goodwin R.M. Model rastu // Socializmus a rast. Cambridge: University Press, 1967.
12. Malkov S.Yu. Sociálna sebaorganizácia a historický proces. Kapitola 2. M.: URSS, 2009.
13. Lanchester F W. Lietadlo vo vojne: Dolná časť štvrtého ramena. Londýn, Constable, 1916.
14. Osipov M.O. O vplyve počtu síl vstupujúcich do boja na ich straty // Vojenská zbierka, jún-október 1915.
15. Bell G. Prey - predátorské rovnice simulujúce imunitnú odpoveď // Math. Biosci. 1973. Číslo 16. S. 291.
16. Marchuk G.I. Matematické modely v imunológii a medicíne. Kapitola 2. M.: Nauka, 1985.
17. Kermack W.O. a Mc.Kendrick A.G. Príspevok k matematickej teórii epidémií // Proceedings of Royal Statistical Society A. 1927. Vol. 115. S. 700.
18. Rabinovič M.I., Muezinolu M.K. Nelineárna dynamika mozgu: emócie a intelektuálna aktivita // UFN. 2010. Číslo 4.
19. Neimark Yu.I. Matematické modely v prírodných vedách a technike. N. Novgorod: Univerzitné vydavateľstvo Nižný Novgorod, 2004.
Saratovský štát prijatý 2.11.2011
Univerzita pomenovaná po N.G. Černyševskij
FENOMÉN MATEMATICKÉHO MODELU LOTKA-VOLTERRA
A PODOBNÉ MODELY
Matematický model Lotka-Volterra (často nazývaný model „predátor-korisť“) je použiteľný na popis rôznych procesov v biológii, ekológii, medicíne, vo výskumoch sociológie, v histórii, rádiofyzike atď.
Varianty tohto modelu sa v tomto prehľade zvažujú metodologicky.
Nasledujúce modely sú sledované: model kontaminácie alebo inej špinavej interakcie s okolím; model triedneho boja; model beztriednej spoločnosti - epock lovcov-zberačov; model vojenských operácií; model vírusových infekčných chorôb; model šírenia epidémie, teda šírenie počítačovej infekcie; model kognitívnych a (alebo) emocionálnych cerebrálnych režimov.
Kľúčové slová: matematický model Lotka-Volterra, systém „predátor-korisť“, analýza prírodných alebo spoločenských javov.
Trubetskov Dmitrij Ivanovič - narodený v Saratove (1938). Absolvent Fyzikálnej fakulty Saratovskej štátnej univerzity (1960). Obhájil dizertačné práce na vedeckú hodnosť kandidáta (1965) a doktora fyzikálnych a matematických vied na SSU (1978) v odbore rádiofyzika. Vedúci Katedry elektroniky, oscilácií a vĺn Fakulty nelineárnych procesov SSU, profesor, člen korešpondent Ruskej akadémie vied, ctený vedec Ruskej federácie, laureát prezidentskej ceny za vzdelávanie. Vedecký riaditeľ Lýcea aplikovaných vied a Fakulty nelineárnych procesov SSU Oblasť vedeckého záujmu: vákuová elektronika a mikrovlnná mikroelektronika, teória kmitov a vĺn, nelineárna dynamika, história vedy. Autor viac ako dvadsiatich učebníc a monografií, ako aj viac ako dvesto článkov v periodikách.
410012, Saratov, ul. Astrachanskaja, 83 rokov
Štátna univerzita Saratov pomenovaná po. N.G. Chernyshevsky E-mail: [chránený e-mailom]
Modely interakcie dvoch typov
Volterrove hypotézy. Analógie s chemickou kinetikou. Volterrove modely interakcií. Klasifikácia typov interakcií Konkurencia. Predátor-korisť. Zovšeobecnené modely interakcií druhov . Kolmogorov model. MacArthurov model interakcie medzi dvoma druhmi hmyzu. Parametrický a fázové portréty Bazykinovho systému.
Za zakladateľa modernej matematickej teórie populácií je právom považovaný taliansky matematik Vito Volterra, ktorý vypracoval matematickú teóriu biologických spoločenstiev, ktorej aparátom sú diferenciálne a integrodiferenciálne rovnice.(Vito Volterra. Lecons sur la Theorie Mathematique de la Lutte pour la Vie. Paríž, 1931). V nasledujúcich desaťročiach sa populačná dynamika vyvíjala najmä v súlade s myšlienkami vyjadrenými v tejto knihe. Ruský preklad Volterrovej knihy vyšiel v roku 1976 pod názvom: „Matematická teória boja o existenciu“ s doslovom Yu.M. Svirezhev, ktorý skúma históriu vývoja matematickej ekológie v období rokov 1931–1976.
Volterrova kniha je napísaná tak, ako sa píšu knihy o matematike. Najprv sformuluje niektoré predpoklady o matematických objektoch, ktoré sa majú študovať, a potom vykoná matematickú štúdiu vlastností týchto objektov.
Systémy, ktoré študoval Volterra, pozostávajú z dvoch alebo viacerých typov. V niektorých prípadoch sa uvažuje o dodávke použitých potravín. Rovnice popisujúce interakciu týchto typov sú založené na nasledujúcich konceptoch.
Volterrove hypotézy
1. Potraviny sú buď dostupné v neobmedzenom množstve, alebo je ich prísun v čase prísne regulovaný.
2. Jedince každého druhu vymierajú tak, že za jednotku času umiera konštantný podiel existujúcich jedincov.
3. Dravé druhy jedia obete a za jednotku času je počet zožratých obetí vždy úmerný pravdepodobnosti stretnutia s jedincami týchto dvoch druhov, t.j. súčin počtu predátorov a počtu koristi.
4. Ak existuje potrava v obmedzenom množstve a niekoľko druhov, ktoré ju dokážu skonzumovať, potom podiel potravy skonzumovanej druhom za jednotku času je úmerný počtu jedincov tohto druhu, pričom sa berie s určitým koeficientom v závislosti od druhov (modely medzidruhovej konkurencie).
5. Ak sa druh živí potravou dostupnou v neobmedzenom množstve, nárast počtu druhov za jednotku času je úmerný počtu druhov.
6. Ak sa druh živí potravou dostupnou v obmedzenom množstve, potom je jeho rozmnožovanie regulované rýchlosťou spotreby potravy, t.j. za jednotku času je nárast úmerný množstvu zjedenej potravy.
Analógie s chemickou kinetikou
Tieto hypotézy majú úzke paralely s chemickou kinetikou. V rovniciach populačnej dynamiky, rovnako ako v rovniciach chemickej kinetiky, sa používa „princíp kolízie“, kedy je rýchlosť reakcie úmerná súčinu koncentrácií reagujúcich zložiek.
V skutočnosti, podľa Volterrových hypotéz, rýchlosť proces Vyhynutie každého druhu je úmerné počtu druhov. V chemickej kinetike to zodpovedá monomolekulárnej reakcii rozkladu určitej látky a v matematickom modeli to zodpovedá záporným lineárnym členom na pravej strane rovníc.
Podľa koncepcií chemickej kinetiky je rýchlosť bimolekulárnej reakcie interakcie medzi dvoma látkami úmerná pravdepodobnosti zrážky týchto látok, t.j. produkt ich koncentrácie. Rovnakým spôsobom je v súlade s Volterrovými hypotézami miera rozmnožovania predátorov (úhyn koristi) úmerná pravdepodobnosti stretnutia medzi predátorom a korisťou, t.j. súčin ich počtu. V oboch prípadoch sa bilineárne členy objavujú v modelovom systéme na pravej strane zodpovedajúcich rovníc.
Nakoniec, lineárne kladné členy na pravej strane Volterrových rovníc, zodpovedajúce rastu populácií za neobmedzených podmienok, zodpovedajú autokatalytickým členom chemických reakcií. Táto podobnosť rovníc v chemických a environmentálnych modeloch nám umožňuje aplikovať rovnaké výskumné metódy pre matematické modelovanie populačnej kinetiky ako pre systémy chemických reakcií.
Klasifikácia typov interakcií
V súlade s Volterrovými hypotézami interakcia dvoch druhov, ktorých počty X 1 a X 2 možno opísať rovnicami:
(9.1)
Tu sú parametre a i - konštanty vlastnej rýchlosti rastu druhu, c ja- konštanty sebaobmedzovania čísel (vnútrodruhová konkurencia), b ij- konštanty interakcie druhov, (i, j= 1,2). Znamienka týchto koeficientov určujú typ interakcie.
V biologickej literatúre sa interakcie zvyčajne klasifikujú podľa príslušných mechanizmov. Rozmanitosť je tu obrovská: rôzne trofické interakcie, chemické interakcie medzi baktériami a planktónnymi riasami, interakcie húb s inými organizmami, postupnosť rastlinných organizmov spojená najmä so súťažou o slnečné svetlo a s vývojom pôd atď. Táto klasifikácia sa zdá byť rozsiahla.
E . Odum, berúc do úvahy modely navrhnuté V. Volterrom, navrhol klasifikáciu nie podľa mechanizmov, ale podľa výsledkov. Podľa tejto klasifikácie by sa vzťahy mali hodnotiť ako pozitívne, negatívne alebo neutrálne v závislosti od toho, či sa početnosť jedného druhu zvyšuje, znižuje alebo zostáva nezmenená v prítomnosti iného druhu. Potom môžu byť hlavné typy interakcií prezentované vo forme tabuľky.
TYPY INTERAKCIE DRUHOV
|
SYMBIÓZA |
b 12 ,b 21 >0 |
||
|
KOMENZALIZMUS |
b 12 ,>0, b 21 =0 |
||
|
DRAVEC-OBETÍ |
b 12 ,>0, b 21 <0 |
||
|
AMENSALIZMUS |
b 12 ,=0, b 21 <0 |
||
|
SÚŤAŽ |
b 12 , b 21 <0 |
||
|
NEUTRALIZMUS |
b 12 , b 21 =0 |
Posledný stĺpec ukazuje znamienka interakčných koeficientov zo systému (9.1)
Pozrime sa na hlavné typy interakcií
SÚŤAŽNÉ ROVNICE:
Ako sme videli v 6. prednáške, súťažné rovnice sú:
 (9.2)
(9.2)
Stacionárne systémové riešenia:
(1).
![]()
Počiatok súradníc pre akékoľvek systémové parametre je nestabilný uzol.
(2).
![]() (9.3)
(9.3)
C stacionárny stav (9.3) je sedlo pri a 1 >b 12 /S 2 a
stabilný uzol pri a 1 12 /s 2 . Táto podmienka znamená, že druh vyhynie, ak je jeho rýchlosť rastu nižšia ako určitá kritická hodnota.
(3).
![]() (9.4)
(9.4)
C stacionárne riešenie (9.4)¾ sedlo pri a 2 >b 21 /c 1 a stabilný uzol na a 2< b 21 /c 1
(4).
![]() (9.5)
(9.5)
Stacionárny stav (9.5) charakterizuje koexistenciu dvoch konkurenčných druhov a predstavuje stabilný uzol, ak je splnený vzťah:
![]()
To znamená nerovnosť:
b 12
b 21
čo nám umožňuje formulovať podmienku koexistencie druhov:
Súčin koeficientov medzipopulačnej interakcie je menší ako súčin koeficientov v rámci populačnej interakcie.
Skutočne, nechajme prirodzené rýchlosti rastu týchto dvoch druhova 1 , a 2 sú rovnaké. Potom bude nevyhnutnou podmienkou stability
c 2 > b 12 ,c 1 > b 21 .
Tieto nerovnosti ukazujú, že nárast veľkosti jedného konkurenta potláča jeho vlastný rast viac ako rast iného konkurenta. Ak sú počty oboch druhov obmedzené, čiastočne alebo úplne, rôznymi zdrojmi, platia vyššie uvedené nerovnosti. Ak majú oba druhy presne rovnaké potreby, potom jeden z nich bude životaschopnejší a vytlačí svojho konkurenta.
Správanie fázových trajektórií systému dáva jasnú predstavu o možných výsledkoch súťaže. Prirovnajme pravé strany rovníc systému (9.2) k nule:
X 1 (a 1 – c 1 X 1 – b 12 X 2) = 0 (dx 1 /dt = 0),
X 2 (a 2 –b 21 X 1 – c 2 X 2) = 0 (dx 2 /dt = 0),
V tomto prípade získame rovnice pre hlavné izokliny systému
X 2 = – b 21 X 1 / c 2 +a 2 /c 2, X 2 = 0
– rovnice izoklinál vertikálnych dotyčníc.
X 2 = – c 1 X 1 / b 12 + a 1 /b 12 , X 1 = 0
– rovnice izoklinál vertikálnych dotyčníc. Body párového priesečníka izoklinál vertikálnych a horizontálnych dotyčnicových sústav predstavujú stacionárne riešenia sústavy rovníc (9.2.), a ich súradnice ![]() sú stacionárne počty konkurenčných druhov.
sú stacionárne počty konkurenčných druhov.
Možné umiestnenie hlavných izoklinál v systéme (9.2) je znázornené na obr. 9.1. Ryža. 9.1Azodpovedá prežitiu druhuX 1, obr. 9.1 b– prežitie druhuX 2, obr. 9.1 V– koexistencia druhov, keď je splnená podmienka (9.6). Obrázok 9.1Gdemonštruje spúšťací systém. Tu závisí výsledok súťaže od počiatočných podmienok. Nenulový stacionárny stav (9.5) pre oba typy je nestabilný. Toto je sedlo, cez ktoré prechádza separatrix a oddeľuje oblasti prežitia každého druhu.

Ryža. 9.1.Umiestnenie hlavných izoklinál na fázovom portréte Volterrovho systému konkurencie dvoch typov (9.2) s rôznymi pomermi parametrov. Vysvetlivky v texte.
Na štúdium konkurencie druhov sa uskutočnili experimenty na širokej škále organizmov. Typicky sa vyberú dva blízko príbuzné druhy a pestujú sa spolu a oddelene za prísne kontrolovaných podmienok. V určitých intervaloch sa vykonáva úplné alebo výberové sčítanie obyvateľstva. Údaje z niekoľkých opakovaných experimentov sa zaznamenávajú a analyzujú. Štúdie sa uskutočnili na prvokoch (najmä nálevniciach), mnohých druhoch chrobákov rodu Tribolium, drosophila a sladkovodných kôrovcoch (dafnie). Na mikrobiálnych populáciách sa uskutočnilo mnoho experimentov (pozri prednášku 11). Pokusy sa uskutočňovali aj v prírode vrátane planárov (Reynolds), dvoch druhov mravcov (Pontin) atď. Na obr. 9.2. znázorňuje rastové krivky rozsievok využívajúcich rovnaký zdroj (zaberajúce rovnakú ekologickú niku). Pri pestovaní v monokultúre Asterionella Formosa dosahuje konštantnú úroveň hustoty a udržuje koncentráciu zdroja (silikátu) na konštantne nízkej úrovni. B. Pri pestovaní v monokultúre Synedrauina chová sa podobným spôsobom a udržuje koncentráciu kremičitanov na ešte nižšej úrovni. B. Počas spoločnej kultivácie (v duplikáte) Synedrauina vytláča Asterionella formosa. Zrejme Synedra

Ryža. 9.2.Konkurencia v rozsievkách. A - pri pestovaní v monokultúre Asterionella Formosa dosahuje konštantnú úroveň hustoty a udržuje koncentráciu zdroja (silikátu) na konštantne nízkej úrovni. b - pri pestovaní v monokultúre Synedrauina chová sa podobným spôsobom a udržuje koncentráciu kremičitanov na ešte nižšej úrovni. V - s kokultiváciou (v duplikáte) Synedruina vytláča Asterionella formosa. Zrejme Synedra vyhráva súťaž vďaka svojej schopnosti plnšie využiť substrát (pozri tiež prednášku 11).
Experimenty G. Gausea o štúdiu konkurencie sú všeobecne známe, demonštrujú prežitie jedného z konkurenčných druhov a umožňujú mu sformulovať „zákon konkurenčného vylúčenia“. Zákon hovorí, že v jednej ekologickej nike môže existovať len jeden druh. Na obr. 9.3. Výsledky Gauseho experimentov sú prezentované pre dva druhy Parametium, ktoré zaberajú rovnakú ekologickú niku (obr. 9.3 a, b) a druhy zaberajúce rôzne ekologické niky (obr. 9.3 c).
Ryža. 9.3. A- Krivky rastu populácie dvoch druhov Parametium v jednodruhových plodinách. Čierne kruhy - P Aurelia, biele kruhy – P. Caudatum
b- Rastové krivky P Aurelia a P . Caudatum v zmiešanej kultúre.
Gause, 1934
Konkurenčný model (9.2) má nevýhody, najmä z neho vyplýva, že koexistencia dvoch druhov je možná len vtedy, ak je ich počet obmedzený rôznymi faktormi, ale model neudáva, aké veľké rozdiely musia byť, aby sa zabezpečilo dlhodobé spolužitie. . Zároveň je známe, že pre dlhodobé spolužitie v meniacom sa prostredí je nevyhnutný rozdiel dosahujúci určitú veľkosť. Zavedenie stochastických prvkov do modelu (napríklad zavedenie funkcie využívania zdrojov) nám umožňuje kvantitatívne skúmať tieto problémy.
Systém PREDATOR+VICTIM
 (9.7)
(9.7)
Tu sú na rozdiel od (9.2) znaky b 12 A b 21 sú odlišné. Rovnako ako v prípade konkurencie, pôvod
![]() (9.8)
(9.8)
je špeciálny bod typu nestabilného uzla. Tri ďalšie možné ustálené stavy:
![]() ,(9.9)
,(9.9)
![]() (9.10)
(9.10)
![]() (9.11)
(9.11)
Je teda možné, aby prežila len korisť (9.10), iba predátor (9.9) (ak má iné zdroje potravy) a koexistencia oboch druhov (9.11). Poslednú možnosť sme už rozoberali v prednáške 5. Možné typy fázových portrétov pre systém dravec-korisť uvádza obr. 9.4.
Izokliny horizontálnych dotyčníc sú priame čiary
X 2 = – b 21 X 1 /c 2 + a 1/c 2, X 2 = 0,
a izokliny vertikálnych dotyčníc– rovný
X 2 = – c 1 X 1 /b 12 + a 2 /b 12 , X 1 = 0.
Stacionárne body ležia na priesečníku zvislých a vodorovných dotyčnicových izoklinál.

Z obr. 9.4 je viditeľné nasledovné. Systém dravec-korisť (9.7) môže mať stabilnú rovnovážnu polohu, v ktorej o Rumová populácia obetí úplne vymrela ( ) a zostali len predátori (obdobie 2 na obr. 9.4 A). Je zrejmé, že takúto situáciu možno realizovať len vtedy, ak okrem typu obetí, o ktoré ide, X 1 dravec X 2 – má prídavné zdroje energie. Táto skutočnosť sa v modeli odráža kladným členom na pravej strane rovnice pre x2. Špeciálne body(1) a (3) (obr. 9.4 A) sú nestabilné. Druhá možnosť – stabilný stacionárny stav, v ktorom populácia predátorov úplne vymrela a zostala len korisť – stabilný bod(3) (obr. 9.4 6 ). Toto je špeciálny bod (1) – tiež nestabilný uzol.
Nakoniec tretia možnosť – udržateľné spolužitie populácií predátorov a koristi (obr. 9.4 V), ktorých stacionárne čísla sú vyjadrené vzorcami (9.11).
Rovnako ako v prípade jednej populácie (pozri prednášku 3), pre model (9.7) Je možné vyvinúť stochastický model, ale nie je možné ho explicitne vyriešiť. Preto sa obmedzíme na všeobecné úvahy. Predpokladajme napríklad, že rovnovážny bod sa nachádza v určitej vzdialenosti od každej z osí. Potom pre fázové trajektórie, na ktorých sú hodnotyX 1 , X 2 dostatočne veľké, deterministický model bude celkom uspokojivý. Ak však v určitom bode fázovej trajektórie nie je akákoľvek premenná príliš veľká, náhodné fluktuácie sa môžu stať významnými. Vedú k tomu, že reprezentujúci bod sa presunie do jednej z osí, čo znamená vyhynutie zodpovedajúceho druhu.
Stochastický model sa teda ukazuje ako nestabilný, pretože stochastický „drift“ skôr či neskôr vedie k vyhynutiu jedného z druhov. V tomto type modelu predátor nakoniec vyhynie, buď náhodou, alebo preto, že jeho populácia koristi je zlikvidovaná ako prvá. Stochastický model systému dravec-korisť dobre vysvetľuje Gauseove experimenty (Gause, 1934), v ktorých riasinkách Paramettum candatum slúžil ako obeť pre iného nálevníka Didinium nasatum – predátor. Očakávané podľa deterministických rovníc (9.7) rovnovážne čísla v týchto pokusoch boli približne len päť jedincov z každého druhu, takže nie je prekvapujúce, že pri každom opakovanom pokuse pomerne rýchlo vymreli buď predátori alebo korisť (a po nich predátori).Výsledky pokusov sú prezentované na obr. 9.5.

Ryža. 9.5. Výška Parametium caudatum a dravé nálevníky Dadinium nasutum. Od : Gause G.F. Boj o existenciu. Baltimore, 1934
Takže analýza Volterrových modelov interakcie druhov ukazuje, že napriek širokej škále typov správania takýchto systémov nemôže v modeli konkurenčných druhov vôbec dochádzať k netlmeným výkyvom v počtoch. Takéto oscilácie sú však pozorované v prírode a pri pokusoch. Potreba ich teoretického vysvetlenia bola jedným z dôvodov formulovania popisov modelov vo všeobecnejšej podobe.
Zovšeobecnené modely interakcie dvoch typov
Na opis interakcie druhov bolo navrhnutých veľké množstvo modelov, ktorých pravé strany rovníc boli funkciami počtu interagujúcich populácií. Bola vyriešená otázka vývoja všeobecných kritérií na stanovenie toho, aký typ funkcií môže opísať správanie dočasnej veľkosti populácie vrátane stabilných fluktuácií. Najznámejšie z týchto modelov patria Kolmogorovovi (1935, revidovaný článok - 1972) a Rosenzweigovi (1963).
 (9.12)
(9.12)
Model obsahuje nasledujúce predpoklady:
1) Dravce medzi sebou neinteragujú, t.j. miera reprodukcie predátorov k 2 a počet obetí L vyhubený za jednotku času jedným predátorom nezávisí od r.
2) Zvýšenie počtu koristi v prítomnosti predátorov sa rovná zvýšeniu neprítomnosti predátorov mínus počet koristi vyhubených predátormi. Funkcie k 1 (X), k 2 (X), L(X), sú spojité a definované na kladnej poloosi X, r³ 0.
3) nevie 1 /dx< 0. To znamená, že miera rozmnožovania koristi v neprítomnosti predátora monotónne klesá s nárastom počtu koristi, čo odráža obmedzenú dostupnosť potravy a iných zdrojov.
4) nevie 2 /dx> 0, k 2 (0) < 0 < k 2 (¥ ). S nárastom počtu obetí sa reprodukčný koeficient predátorov monotónne znižuje s nárastom počtu obetí, pričom sa pohybuje od negatívnych hodnôt (keď nie je čo jesť) k pozitívnym.
5) Počet zlikvidovanej koristi jedným predátorom za jednotku času L(X)> 0 pri N> 0; L(0)=0.
Možné typy fázových portrétov systému (9.12) sú uvedené na obr. 9.6:

Ryža. 9.6.Fázové portréty systému Kolmogorov (9.12), ktorý popisuje interakciu dvoch typov pri rôznych pomeroch parametrov. Vysvetlivky v texte.
Stacionárne riešenia (sú dve alebo tri) majú tieto súradnice:
(1). ` x=0;` y=0.
Počiatok súradníc pre ľubovoľné hodnoty parametrov je sedlo (obr. 9.6 a-d).
(2). ` x=A,` y=0.(9.13)
Aurčí sa z rovnice:
k 1 (A)=0.
Stacionárne riešením (9.13) je sedlo ak B< A (Obr. 9.6 A, b, G), B určené z rovnice
k 2 (B)=0
Bod (9.13) sa umiestni do kladného kvadrantu, ak B>A . Toto je stabilný uzol .
Posledný prípad, ktorý zodpovedá úhynu dravca a prežitiu koristi, je znázornený na obr. 9.6 V.
(3). ` x=B,` y=C.(9.14)
Hodnota C sa určí z rovníc:
Bod (9.14) – ohnisko (obr.9.6 A) alebo uzol (obr.9.6 G), ktorej stabilita závisí od znamienka veličinys
s 2 = – k 1 (B) – k 1 (B)B+L(B)C.
Ak s>0, bod je stabilný, aks<0 ‑ точка неустойчива, и вокруг нее могут существовать предельные циклы (рис. 9.6 b)
V zahraničnej literatúre sa častejšie uvažuje o podobnom modeli, ktorý navrhli Rosenzweig a MacArthur (1963):
 (9.15)
(9.15)
Kde f(X) - miera zmeny počtu obetí X v neprítomnosti predátorov, F( x, y) - intenzita predácie, k- koeficient charakterizujúci účinnosť spracovania biomasy koristi na biomasu predátorov, e- úmrtnosť predátorov.
Model (9.15) sa redukuje na špeciálny prípad Kolmogorovovho modelu (9.12) za nasledujúcich predpokladov:
1) počet predátorov je obmedzený iba počtom koristi,
2) rýchlosť, akou daný dravec požiera korisť, závisí len od hustoty populácie koristi a nezávisí od hustoty populácie dravca.
Potom nadobudnú tvar rovnice (9.15).

Pri popise interakcie skutočných druhov sú pravé strany rovníc špecifikované v súlade s predstavami o biologickej realite. Zoberme si jeden z najpopulárnejších modelov tohto typu.
Model interakcie medzi dvoma druhmi hmyzu (MacArthur, 1971)
Model, ktorý budeme uvažovať nižšie, bol použitý na vyriešenie praktického problému kontroly škodlivého hmyzu sterilizáciou samcov jedného z druhov. Na základe biologických vlastností interakcie druhov bol napísaný nasledujúci model
 (9.16)
(9.16)
Tu x, y- biomasa dvoch druhov hmyzu. Trofické interakcie druhov opísaných v tomto modeli sú veľmi zložité. Toto určuje tvar polynómov na pravej strane rovníc.
Pozrime sa na pravú stranu prvej rovnice. Druhy hmyzu X jesť larvy druhu pri(člen +k 3 y), ale dospelí druhu pri jesť larvy druhu X podlieha vysokej početnosti druhov X alebo pri alebo oba typy (členovia –k 4 xy, – y 2). Pri malom Xúmrtnosť druhov X vyšší ako jeho prirodzený prírastok (1 –k 1 +k 2 x–x 2 < 0 pri malom X). V druhej rovnici člen k 5 odráža prirodzený rast druhu y; –k 6 y – sebaovládanie tohto typu,–k 7 X– požieranie lariev druhu pri druhy hmyzu x, k 8 xy – zvýšenie biomasy druhov pri v dôsledku konzumácie dospelým hmyzom tohto druhu pri larvy druhu X.
Na obr. 9.7 je prezentovaný limitný cyklus, ktorý je trajektóriou stabilného periodického riešenia sústavy (9.16).

Riešenie otázky, ako zabezpečiť koexistenciu populácie s jej biologickým prostredím, samozrejme nemožno získať bez zohľadnenia špecifík konkrétneho biologického systému a rozboru všetkých jeho vzájomných vzťahov. Štúdium formálnych matematických modelov nám zároveň umožňuje odpovedať na niektoré všeobecné otázky. Možno tvrdiť, že pre modely ako (9.12) skutočnosť kompatibility alebo nekompatibility populácií nezávisí od ich počiatočnej veľkosti, ale je určená iba povahou interakcie druhov. Model pomáha odpovedať na otázku: ako ovplyvniť biocenózu a riadiť ju, aby sa škodlivé druhy rýchlo zničili.
Manažment možno zredukovať na krátkodobú, náhlu zmenu hodnôt populácie X A u. Táto metóda zodpovedá kontrolným metódam, ako je jednorazové zničenie jednej alebo oboch populácií chemickými prostriedkami. Z vyššie formulovaného tvrdenia je zrejmé, že pre kompatibilné populácie bude tento spôsob kontroly neúčinný, keďže systém časom opäť dosiahne stacionárny režim.
Ďalším spôsobom je zmena typu interakčných funkcií medzi zobrazeniami, napríklad pri zmene hodnôt systémových parametrov. Práve tejto parametrickej metóde zodpovedajú metódy biologickej kontroly. Pri zavádzaní sterilizovaných samcov sa teda tempo prirodzeného rastu populácie znižuje. Ak zároveň získame iný typ fázového portrétu, taký, kde je iba stabilný stacionárny stav s nulovým počtom škodcov, kontrola povedie k požadovanému výsledku. – ničenie populácie škodlivého druhu. Je zaujímavé poznamenať, že niekedy je vhodné aplikovať zásah nie na samotného škodcu, ale na jeho partnera. Vo všeobecnosti sa nedá povedať, ktorá metóda je účinnejšia. To závisí od dostupných ovládacích prvkov a od explicitnej formy funkcií opisujúcich interakciu populácií.
Model od A.D. Bazykina
Teoretická analýza modelov interakcií druhov bola najkomplexnejšie vykonaná v knihe A.D. Bazykina „Biophysics of Interacting Populations“ (M., Nauka, 1985).
Uvažujme o jednom z modelov predátor-korisť študovaných v tejto knihe.
 (9.17)
(9.17)
Systém (9.17) je zovšeobecnením najjednoduchšieho modelu Volterra predátor-korisť (5.17) s prihliadnutím na vplyv saturácie predátora. Model (5.17) predpokladá, že intenzita pasenia koristi sa lineárne zvyšuje so zvyšujúcou sa hustotou koristi, čo nezodpovedá realite pri vysokých hustotách koristi. Na opis závislosti stravy predátora od hustoty koristi možno zvoliť rôzne funkcie. Najdôležitejšie je, aby zvolená funkcia s rastom X inklinoval asymptoticky ku konštantnej hodnote. Model (9.6) využíval logistickú závislosť. V Bazykinovom modeli je ako taká funkcia zvolená hyperbola X/(1+px). Pripomeňme si, že ide o formu Monodovho vzorca, ktorý popisuje závislosť rýchlosti rastu mikroorganizmov od koncentrácie substrátu. Korisť tu hrá úlohu substrátu a predátor úlohu mikroorganizmov. .
Systém (9.17) závisí od siedmich parametrov. Počet parametrov možno znížiť nahradením premenných:
X® (A/D)X; r ® (A/D)/y;
t® (1/A)t; g (9,18)
a závisí od štyroch parametrov.
Pre kompletnú kvalitatívnu štúdiu je potrebné rozdeliť priestor štvorrozmerných parametrov na oblasti s rôznymi typmi dynamického správania, t.j. vytvoriť parametrický alebo štrukturálny portrét systému.
Potom je potrebné zostrojiť fázové portréty pre každú z oblastí parametrického portrétu a opísať bifurkácie, ktoré sa vyskytujú pri fázových portrétoch na hraniciach rôznych oblastí parametrického portrétu.
Konštrukcia kompletného parametrického portrétu sa vykonáva vo forme sady „rezov“ (projekcií) nízkorozmerného parametrického portrétu s pevnými hodnotami niektorých parametrov.
Parametrický portrét systému (9.18) pre pevné g a malé e znázornené na obr. 9.8. Portrét obsahuje 10 oblastí s rôznymi typmi správania fázových trajektórií.
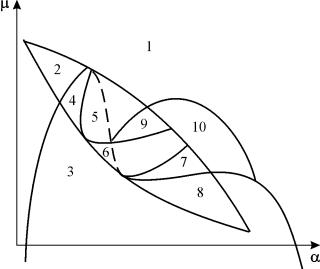
Ryža. 9.8.Parametrický portrét systému (9.18) pre pevnég
a malé e
Správanie systému pri rôznych pomeroch parametrov môže byť výrazne odlišné (obr. 9.9). Systém umožňuje:
1) jedna stabilná rovnováha (oblasti 1 a 5);
2) jeden stabilný limitný cyklus (regióny 3 a 8);
3) dve stabilné rovnováhy (región 2)
4) stabilný limitný cyklus a nestabilná rovnováha v ňom (oblasti 6, 7, 9, 10)
5) stabilný limitný cyklus a stabilná rovnováha mimo neho (región 4).
V parametrických oblastiach 7, 9, 10 je oblasť príťažlivosti rovnováhy obmedzená nestabilným limitným cyklom ležiacim vo vnútri stabilného. Najzaujímavejšou štruktúrou je fázový portrét, ktorý zodpovedá oblasti 6 v parametrickom portréte. Podrobne je to znázornené na obr. 9.10.

Oblasť príťažlivosti rovnováhy B 2 (vytieňovaná) je „slimák“ krútiaci sa z nestabilného ohniska B 1. Ak je známe, že v počiatočnom okamihu bol systém v blízkosti B 1, potom je možné posúdiť, či zodpovedajúca trajektória dosiahne rovnováhu B 2 alebo stabilný limitný cyklus obklopujúci tri rovnovážne body C (sedlo), B 1 a B 2 na základe pravdepodobnostných úvah.

Obr.9.10.Fázový portrét systému 9.18 pre parametrickú oblasť 6. Oblasť príťažlivosti B 2 je zatienená
V parametrickom portréte(9.7) je ich 22 rôzne bifurkačné hranice, ktoré sa tvoria 7 rôzne typy bifurkácií. Ich štúdium nám umožňuje identifikovať možné typy správania systému pri zmene jeho parametrov. Napríklad pri presune z oblasti 1 do oblasti 3 nastáva zrod malého limitného cyklu alebo mäkké zrodenie vlastných oscilácií okolo jedinej rovnováhy IN. Podobné mäkké zrodenie vlastných oscilácií, ale okolo jednej z rovnováh, a to B 1 , dochádza pri prekračovaní hraníc regiónov 2 a 4. Pri odchode z areálu 4 do oblasti 5 stabilný limitný cyklus okolo boduB 1 „praskne“ na slučke oddeľovačov a jediným priťahujúcim bodom zostáva rovnováha B 2 atď.
Pre prax je, samozrejme, mimoriadne zaujímavý vývoj kritérií pre blízkosť systému k hraniciam rozvetvenia. Biológovia sú si skutočne dobre vedomí vlastnosti „tlmenia“ alebo „pružnosti“ prírodných ekologických systémov. Tieto výrazy zvyčajne označujú schopnosť systému absorbovať vonkajšie vplyvy. Pokiaľ intenzita vonkajšieho vplyvu nepresiahne určitú kritickú hodnotu, nedochádza v správaní systému ku kvalitatívnym zmenám. Vo fázovej rovine to zodpovedá návratu sústavy do stabilného stavu rovnováhy alebo do stabilného limitného cyklu, ktorého parametre sa od pôvodného príliš nelíšia. Keď intenzita nárazu prekročí povolenú úroveň, systém sa „rozpadne“ a prejde do kvalitatívne iného režimu dynamického správania, napríklad jednoducho vyhasne. Tento jav zodpovedá prechodu bifurkácie.
Každý typ bifurkačného prechodu má svoje charakteristické črty, ktoré umožňujú posúdiť nebezpečenstvo takéhoto prechodu pre ekosystém. Tu je niekoľko všeobecných kritérií označujúcich blízkosť nebezpečnej hranice. Rovnako ako v prípade jedného druhu, ak sa pri znížení počtu jedného z druhov systém „zasekne“ v blízkosti nestabilného sedlového bodu, čo sa prejaví veľmi pomalým obnovením počtu na pôvodnú hodnotu, potom systém je blízko kritickej hranice. Indikátorom nebezpečenstva je aj zmena tvaru kolísania počtu dravcov a koristi. Ak sa oscilácie, ktoré sú blízke harmonickým, stanú relaxačnými a amplitúda oscilácií sa zvýši, môže to viesť k strate stability systému a vyhynutiu jedného z druhov.
Ďalšie prehlbovanie matematickej teórie interakcie medzi druhmi ide v línii podrobností o štruktúre samotných populácií a pri zohľadnení časových a priestorových faktorov.
Literatúra.
Kolmogorov A.N. Kvalitatívna štúdia matematických modelov populačnej dynamiky. // Problémy kybernetiky. M., 1972, číslo 5.
MacArtur R. Grafická analýza ekologických systémov // Správa oddelenia biológie Perinceton University. 1971
A.D. Bazykin „Biofyzika interagujúcich populácií“. M., Nauka, 1985.
V. Volterra: „Matematická teória boja o existenciu“. M.. Veda, 1976
Gause G.F. Boj o existenciu. Baltimore, 1934.
Populačná dynamika je jednou z oblastí matematického modelovania. Je zaujímavý, pretože má špecifické aplikácie v biológii, ekológii, demografii a ekonómii. V tejto časti je niekoľko základných modelov, z ktorých jeden, model „Predator-Prey“, je popísaný v tomto článku.
Prvým príkladom modelu v matematickej ekológii bol model navrhnutý V. Volterrom. Bol to on, kto prvý zvážil model vzťahu medzi predátorom a korisťou.
Pozrime sa na vyhlásenie o probléme. Nech sú dva druhy zvierat, z ktorých jedno požiera druhé (dravce a korisť). V tomto prípade sa vychádza z nasledujúcich predpokladov: zdroje potravy koristi nie sú obmedzené, a preto v neprítomnosti predátora sa populácia koristi zvyšuje podľa exponenciálneho zákona, zatiaľ čo predátori, oddelení od svojich obetí, postupne umierajú. hladu, tiež podľa exponenciálneho zákona. Akonáhle začnú predátori a korisť žiť vo vzájomnej tesnej blízkosti, zmeny vo veľkosti ich populácie budú vzájomne prepojené. V tomto prípade bude relatívny nárast počtu koristi samozrejme závisieť od veľkosti populácie predátorov a naopak.
V tomto modeli sa predpokladá, že všetci predátori (a všetka korisť) sú v rovnakých podmienkach. Potravinové zdroje obetí sú zároveň neobmedzené a predátori sa živia výlučne obeťami. Obe populácie žijú na obmedzenom území a neinteragujú so žiadnymi inými populáciami a neexistujú žiadne ďalšie faktory, ktoré by mohli ovplyvniť veľkosť populácie.
Samotný matematický model „predátor-korisť“ pozostáva z dvojice diferenciálnych rovníc, ktoré popisujú dynamiku populácií dravcov a koristi v najjednoduchšom prípade, keď existuje jedna populácia predátorov a jedna populácia koristi. Vzorec je charakterizovaný kolísaním veľkosti oboch populácií, pričom vrchol u predátorov je mierne za vrcholom u koristi. Tento model možno nájsť v mnohých prácach o populačnej dynamike alebo matematickom modelovaní. Bol široko pokrytý a analyzovaný pomocou matematických metód. Vzorce však nemusia vždy poskytnúť jasnú predstavu o prebiehajúcom procese.
Je zaujímavé zistiť, ako presne závisí populačná dynamika v tomto modeli od počiatočných parametrov a nakoľko to zodpovedá realite a zdravému rozumu, a vidieť to graficky bez toho, aby sme sa uchyľovali k zložitým výpočtom. Na tento účel bol na základe modelu Volterra vytvorený program v prostredí Mathcad14.
Najprv skontrolujme, či model vyhovuje reálnym podmienkam. Aby sme to urobili, uvažujme o degenerovaných prípadoch, keď v daných podmienkach žije iba jedna z populácií. Teoreticky sa ukázalo, že pri absencii predátorov sa populácia koristi v čase neobmedzene zvyšuje a populácia predátorov bez koristi vymiera, čo vo všeobecnosti zodpovedá modelu a reálnej situácii (s uvedenou formuláciou tzv. problém).
Získané výsledky odzrkadľujú tie teoretické: predátori postupne vymierajú (obr. 1) a počet koristi sa neobmedzene zvyšuje (obr. 2).
Obr. 1 Závislosť počtu predátorov od času v neprítomnosti koristi

Obr. 2 Závislosť počtu koristi od času v neprítomnosti predátorov
Ako vidno, v týchto prípadoch systém zodpovedá matematickému modelu.
Pozrime sa, ako sa systém správa pri rôznych počiatočných parametroch. Nech sú dve populácie - levy a antilopy - predátori a korisť a sú uvedené počiatočné ukazovatele. Potom dostaneme nasledujúce výsledky (obr. 3):
Tabuľka 1. Koeficienty oscilačného režimu systému

Obr.3 Systém s hodnotami parametrov z tabuľky 1
Analyzujme získané údaje na základe grafov. S počiatočným nárastom populácie antilop sa pozoruje nárast počtu predátorov. Všimnite si, že vrcholný nárast populácie predátorov sa pozoruje neskôr, počas poklesu populácie koristi, čo je celkom v súlade so skutočnými konceptmi a matematickým modelom. Zvýšenie počtu antilop skutočne znamená zvýšenie zdrojov potravy pre levy, čo znamená zvýšenie ich počtu. Ďalej, aktívna konzumácia antilop levmi vedie k rýchlemu poklesu počtu koristi, čo nie je prekvapujúce vzhľadom na chuť predátora, alebo skôr na frekvenciu predátorov, ktorí jedia korisť. Postupné znižovanie počtu predátorov vedie k situácii, keď sa populácia koristi ocitne v podmienkach priaznivých pre rast. Potom sa situácia s určitým obdobím opakuje. Dospeli sme k záveru, že tieto podmienky nie sú vhodné pre harmonický vývoj jedincov, pretože majú za následok prudký pokles populácie koristi a prudký nárast v oboch populáciách.
Nastavme teraz počiatočný počet predátorov na rovných 200 jedincov pri zachovaní ostatných parametrov (obr. 4).
Tabuľka 2. Koeficienty oscilačného režimu systému

Obr.4 Systém s hodnotami parametrov z tabuľky 2
Teraz systém osciluje prirodzenejšie. Za týchto predpokladov systém existuje celkom harmonicky, nedochádza k prudkým nárastom a poklesom počtu v oboch populáciách. Dospeli sme k záveru, že s týmito parametrami sa obe populácie vyvíjajú dostatočne rovnomerne na to, aby žili spolu na rovnakom území.
Počiatočný počet predátorov nastavme na 100 jedincov, počet koristi na 200 pri zachovaní ostatných parametrov (obr. 5).
Tabuľka 3. Koeficienty oscilačného režimu systému

Obr.5 Systém s hodnotami parametrov z tabuľky 3
V tomto prípade je situácia blízka prvej uvažovanej situácii. Všimnite si, že pri vzájomnom náraste populácií sa prechody z nárastu na pokles populácie koristi stali plynulejšími a populácia predátorov zostáva pri absencii koristi na vyššej číselnej hodnote. Dospeli sme k záveru, že keď jedna populácia úzko súvisí s druhou, ich interakcia nastáva harmonickejšie, ak sú špecifické počiatočné populácie dostatočne veľké.
Zvážme zmenu ďalších parametrov systému. Nech počiatočné čísla zodpovedajú druhému prípadu. Zvýšme mieru reprodukcie obetí (obr. 6).
Tabuľka 4. Koeficienty oscilačného režimu systému

Obr.6 Systém s hodnotami parametrov z tabuľky 4
Porovnajme tento výsledok s výsledkom získaným v druhom prípade. V tomto prípade sa pozoruje rýchlejší rast obete. Predátor aj korisť sa v tomto prípade správajú ako v prvom prípade, čo bolo vysvetlené nízkou veľkosťou populácie. S touto interakciou dosahujú obe populácie vrchol pri hodnotách oveľa vyšších ako v druhom prípade.
Teraz zvýšme tempo rastu predátorov (obr. 7).
Tabuľka 5. Koeficienty oscilačného režimu systému

Obr.7 Systém s hodnotami parametrov z tabuľky 5
Rovnakým spôsobom porovnajme výsledky. V tomto prípade zostávajú všeobecné charakteristiky systému rovnaké, s výnimkou zmeny obdobia. Ako sa dalo očakávať, obdobie sa skrátilo, čo sa vysvetľuje rýchlym poklesom populácie predátorov v neprítomnosti koristi.
A nakoniec zmeňme koeficient medzidruhovej interakcie. Po prvé, zvýšime frekvenciu predátorov, ktorí požierajú korisť:
Tabuľka 6. Koeficienty oscilačného režimu systému

Obr.8 Systém s hodnotami parametrov z tabuľky 6
Keďže dravec požiera svoju korisť častejšie, maximálna veľkosť populácie sa v porovnaní s druhým prípadom zväčšila a zmenšil sa aj rozdiel medzi maximálnou a minimálnou veľkosťou populácie. Obdobie oscilácie systému zostáva rovnaké.
A teraz znížme frekvenciu predátorov, ktorí jedia korisť:
Tabuľka 7. Koeficienty oscilačného režimu systému

Obr.9 Systém s hodnotami parametrov z tabuľky 7
Teraz predátor žerie korisť menej často, maximálna veľkosť populácie sa v porovnaní s druhým prípadom znížila a maximálna veľkosť populácie koristi sa zvýšila 10-krát. Z toho vyplýva, že za týchto podmienok má populácia koristi väčšiu slobodu z hľadiska rozmnožovania, pretože dravec potrebuje na jej dostatok menej hmoty. Znížil sa aj rozdiel medzi maximálnou a minimálnou veľkosťou populácie.
Pri pokuse o modelovanie zložitých procesov v prírode alebo spoločnosti tak či onak vyvstáva otázka správnosti modelu. Prirodzene, pri modelovaní sa proces zjednodušuje a niektoré drobné detaily sú zanedbané. Na druhej strane existuje nebezpečenstvo prílišného zjednodušenia modelu, čím sa vymažú dôležité črty javu spolu s nepodstatnými. Aby sa predišlo tejto situácii, pred modelovaním je potrebné preštudovať si predmet, v ktorom sa tento model používa, preskúmať všetky jeho charakteristiky a parametre, a čo je najdôležitejšie, vyzdvihnúť tie vlastnosti, ktoré sú najvýznamnejšie. Proces musí mať prirodzený popis, intuitívne zrozumiteľný, zhodujúci sa v hlavných bodoch s teoretickým modelom.
Model uvažovaný v tejto práci má množstvo významných nevýhod. Napríklad predpoklad neobmedzených zdrojov pre obeť, absencia faktorov tretích strán ovplyvňujúcich úmrtnosť oboch druhov atď. Všetky tieto predpoklady neodrážajú skutočný stav. Napriek všetkým nedostatkom sa však model rozšíril v mnohých oblastiach, dokonca ďaleko od ekológie. To možno vysvetliť skutočnosťou, že systém „predátor-korisť“ poskytuje všeobecnú predstavu o interakcii druhov. Interakcie s prostredím a inými faktormi môžu byť opísané inými modelmi a analyzované spoločne.
Vzťahy typu „predátor – korisť“ sú podstatnou črtou rôznych druhov životných aktivít, pri ktorých dochádza ku kolízii dvoch interagujúcich strán. Tento model sa odohráva nielen v ekológii, ale aj v ekonomike, politike a iných oblastiach činnosti. Napríklad jednou z oblastí súvisiacich s ekonomikou je analýza trhu práce, ktorá zohľadňuje dostupných potenciálnych pracovníkov a voľné pracovné miesta. Táto téma by bola zaujímavým pokračovaním práce na modeli dravec-korisť.
Dovoliť a byť počet koristi a predátorov, resp. Predpokladajme, že jediným limitujúcim faktorom obmedzujúcim rozmnožovanie koristi je tlak na ne zo strany predátorov a rozmnožovanie dravcov je limitované množstvom potravy, ktorú získavajú (množstvom koristi). Potom pri absencii predátorov by mal počet koristi rásť exponenciálne s relatívnou rýchlosťou a a predátori by pri absencii koristi mali tiež exponenciálne vymierať s relatívnou rýchlosťou t. Koeficienty a a m sú koeficienty prirodzeného zvýšenie koristi a prirodzenej úmrtnosti dravcov, resp.
Nech je počet (alebo biomasa) koristi skonzumovanej jedným predátorom za jednotku času a časť energie získanej z tejto biomasy dravec minie na rozmnožovanie a zvyšok minie na udržanie bazálneho metabolizmu a lovecké aktivity. Potom rovnicu systému dravec-korisť možno zapísať ako

Funkcia sa zvyčajne nazýva trofická funkcia dravca alebo funkčná odpoveď (funkčná odpoveď) dravca na hustotu populácie koristi. Práve tieto funkcie sú bežne identifikované v experimentálnych štúdiách predácie a v súčasnosti sa uznáva, že tieto funkcie zvyčajne patria k jednému z nasledujúcich troch typov (obr. 11). Dynamické správanie systému zjavne do značnej miery závisí od typu trofickej funkcie.

Ryža. 11. Rôzne typy trofických funkcií v systéme dravec – korisť: a) tento typ je charakteristický pre bezstavovce a niektoré druhy dravých rýb; b) trofická funkcia s výrazným prahom nasýtenia je charakteristická pre predátorov, ktorí sa živia filtrom (napríklad veľa mäkkýšov); c) tento typ je charakteristický pre stavovce - organizmy, ktoré vykazujú pomerne zložité správanie (napríklad schopné učenia). Trofická funkcia bude mať podobný vzhľad, ak si korisť dokáže vyvinúť obrannú stratégiu (napríklad schovanie sa v úkryte neprístupnom predátorom).
Pre malé hodnoty x, napríklad, keď sú trofické vzťahy v systéme napäté a takmer všetka korisť sa stáva korisťou predátora, ktorý je vždy hladný a nenasýti sa (situácia je v prírode pomerne bežná), trofický funkciu možno považovať za lineárnu funkciu počtu koristi, t.j. Okrem toho predpokladajme, že . Potom

Systém (2.2) sa až do notácie zhoduje s klasickým modelom dravec-korisť V. Volterra, ktorý ukázal, že tento systém má integrál v tvare
![]()
kde Ak sú počiatočné hodnoty počtu koristi a dravcov, potom
![]()
a rovnica (2.3) popisuje rodinu vnorených uzavretých kriviek zodpovedajúcich fázovým trajektóriám periodických riešení systému (2.2).

Ryža. 12. Fázový portrét dravca klasického systému Volterra - korisť.
Všimnite si, že ako sa C zvyšuje, amplitúdy kmitov a y sa zvyšujú (obr. 12). Pri minimálnej hodnote sa tieto krivky stiahnu do bodu so súradnicami . Je ľahké vidieť, že sú to riešenia systému (2.2) pre , t.j. jeho netriviálnu rovnováhu. Pre prípad malých kmitov blízko tohto stavu možno modelové rovnice zapísať ako
![]()
Korene zodpovedajúcej charakteristickej rovnice sú , t.j. bod je stred. Obdobie malých oscilácií, pričom oscilácie v počte jedného druhu sú fázovo posunuté voči osciláciám iného druhu o
V sústave (2.2) je ešte jedna rovnovážna poloha - počiatok. Je ľahké vidieť, že tento bod je sedlom. Súradnicové osi sú oddelené, pričom os vstupuje do sedla a os ho opúšťa (pozri obr. 12).
Napriek tomu, že Volterrov model dokázal vysvetliť mnohé skutočne pozorované javy, má veľkú nevýhodu - nehrubosť (v matematickom zmysle slova) Volterrových cyklov, takže pri akýchkoľvek, akokoľvek slabých, poruchách fázových súradníc, systém prechádza z jedného cyklu do druhého. Túto vlastnosť „drsnosti“ by zrejme mali mať adekvátnejšie modely.
Z hľadiska teórie stability je rovnovážny stav systému (x, y) stavom indiferentnej rovnováhy, stabilný podľa Ljapunova, ale nie asymptoticky.Neprítomnosť asymptotickej stability rovnováhy naznačuje, že vo Volterrovom systéme neexistujú žiadne mechanizmy, ktoré by sa snažili zachovať jej netriviálny rovnovážny stav.
Vyššie uvedené tvrdenie o stabilite rovnováhy je celkom ľahko dokázané zostrojením zodpovedajúcej Ljapunovovej funkcie. Ale o tom viac v ďalšom odseku.

Podobné články



