Още през 20-те години. А. Лотка и малко по-късно, независимо от него, В. Волтер, предложиха математически модели, които описват съчетани колебания в броя на хищниците и жертвите.
Моделът се състои от два компонента:
C – брой хищници; N – брой жертви;
Да приемем, че при отсъствието на хищници популацията на плячката ще нараства експоненциално: dN/dt = rN. Но плячката се унищожава от хищници със скорост, която се определя от честотата на срещи между хищника и плячката, а честотата на срещите се увеличава с увеличаване на броя на хищника (C) и плячката (N). Точният брой на срещаната и успешно изядена плячка ще зависи от ефективността, с която хищникът намира и хваща плячката, т.е. от a’ – „ефективност на търсенето” или „честота на атака”. По този начин честотата на „успешните“ срещи между хищника и жертвата и следователно скоростта на изяждане на жертвите ще бъде равна на a’СN и общо: dN/dt = rN – a’CN (1*).
При липса на храна отделните хищници губят тегло, гладуват и умират. Да приемем, че в разглеждания модел размерът на популацията на хищник при липса на храна поради глад ще намалее експоненциално: dC/dt = - qC, където q е смъртността. Смъртта се компенсира от раждането на нови индивиди със скорост, която според този модел зависи от две обстоятелства:
1) норма на консумация на храна, a’CN;
2) ефективността (f), с която тази храна преминава в потомството на хищника.
Така плодовитостта на хищника е равна на fa’CN и общо: dC/dt = fa’CN – qC (2*). Уравнения 1* и 2* представляват модела на Lotka-Wolter. Свойствата на този модел могат да бъдат изследвани, могат да бъдат конструирани линейни изоклини, които съответстват на постоянен размер на популацията, и с помощта на такива изоклини може да се определи поведението на взаимодействащите популации хищник-плячка.
В случай на популация плячка: dN/dt = 0, rN = a’CN или C = r/a’. защото r и a’ = const, изоклина за жертвата ще бъде линията, за която стойността на C е постоянна:
При ниска плътност на хищника (C), броят на плячката (N) се увеличава, напротив, намалява;
По подобен начин за хищници (уравнение 2*) с dC/dt = 0, fa’CN = qC или N = q/fa’, т.е. Изоклина за хищник ще бъде линия, по която N е постоянна: При висока плътност на плячката, размерът на популацията на хищника се увеличава, а при ниска плътност намалява.
Техният брой претърпява неограничени конюгатни колебания. Когато броят на плячката е голям, броят на хищниците се увеличава, което води до увеличаване на натиска на хищниците върху популацията на плячката и по този начин до намаляване на нейния брой. Това намаление от своя страна води до ограничаване на хищниците в храната и спад в техния брой, което води до отслабване на натиска на хищниците и увеличаване на броя на плячката, което отново води до увеличаване на популацията на хищници и т.н.
Популациите извършват един и същ цикъл от колебания за неопределено време за дълго време, докато някакво външно влияние промени числеността им, след което популациите извършват нови цикли от неограничени колебания. Всъщност околната среда непрекъснато се променя и популациите непрекъснато ще се изместват към нови нива. За да бъдат редовни циклите на колебания, които една популация прави, те трябва да са стабилни: ако външно влияние промени нивото на популацията, тогава те трябва да се стремят към първоначалния цикъл. Такива цикли се наричат стабилни, гранични цикли.
Моделът на Lotka-Wolter ни позволява да покажем основната тенденция във връзката хищник-жертва, която се изразява в появата на флуктуации в популацията на плячката, придружени от флуктуации в популацията на хищника. Основният механизъм на такива флуктуации е забавянето във времето, присъщо на последователността от състояния от голям брой плячка до голям брой хищници, след това към малък брой плячка и малък брой хищници, до голям брой плячка, и т.н.
5) ПОПУЛАЦИОННИ СТРАТЕГИИ НА ХИЩНИЦИ И ПРИМАТИ
Връзката „хищник-плячка“ представлява връзките в процеса на пренос на материя и енергия от фитофаги към зоофаги или от хищници от по-нисък ред към хищници по-висок ред. от Въз основа на характера на тези взаимоотношения се разграничават три вида хищници:
а) събирачи. Хищникът събира малки, доста многобройни подвижни жертви. Този вид хищничество е характерно за много видове птици (пловка, чинка, конник и др.), Които изразходват енергия само за търсене на жертви;
б) истински хищници. Хищникът дебне и убива плячката;
V) скотовъдци. Тези хищници използват плячка многократно, например мухи или конски мухи.
Стратегията за получаване на храна сред хищниците е насочена към осигуряване на енергийна ефективност на храненето: енергийните разходи за получаване на храна трябва да бъдат по-малки от енергията, получена по време на нейното асимилиране.
Истински хищницисе разделят на
„жътвари“, които се хранят с изобилие от ресурси (включително планктонна риба и дори усати китове), и „ловци“, които търсят по-малко изобилна храна. На свой ред
„Ловците“ се делят на „засади“, които чакат плячка (например щука, ястреб, котка, бръмбар богомолка), „търсачи“ (насекомоядни птици) и „преследвачи“. За последната група търсенето на храна не изисква големи разходи на енергия, но много от нея са необходими, за да овладеят плячката (лъвовете в саваните). Някои хищници обаче могат да комбинират елементи от стратегията на различни опции за лов.
Както при връзката „фитофаг-растение“, в природата не се наблюдава ситуация, при която всички жертви са изядени от хищници, което в крайна сметка води до тяхната смърт. Екологичен баланс между хищници и плячка се поддържа чрез специални механизми, намалявайки риска от пълно унищожаване на жертвите. Така че жертвите могат:
Бягайте от хищник. В този случай в резултат на адаптации се увеличава мобилността както на жертвите, така и на хищниците, което е особено характерно за степните животни, които нямат къде да се скрият от преследвачите си;
Придобийте защитно оцветяване („преструвайте се“ на листа или клонки) или, обратно, ярък цвят, N.: червено, предупреждаващо хищниците за горчивия вкус. Добре известно е, че цветът на заека се променя през различните периоди от годината, което му позволява да се маскира в тревата през лятото и на фона на белия сняг през зимата. Адаптивни промени в цвета могат да възникнат на различни етапи от онтогенезата: малките на тюлените са бели (снежен цвят), а възрастните са черни (скалисти брегове);
Разпределят се на групи, което прави търсенето и улавянето им по-енергоемко за хищника;
Скрийте се в убежища;
Преминете към мерки за активна защита (тревопасни животни с рога, бодливи риби), понякога съвместни (мускусните говеда могат да предприемат „пълна защита“ от вълци и др.).
От своя страна хищниците развиват не само способността бързо да преследват плячка, но и обоняние, което им позволява да определят местоположението на плячката по миризмата. Много видове хищници разкъсват дупките на жертвите си (лисици, вълци).
В същото време самите те правят всичко възможно, за да избегнат откриването на тяхното присъствие. Това обяснява чистотата на малките котки, които прекарват много време в тоалетна и заравяне на екскременти, за да премахнат миризмите. Хищниците носят „камуфлажни дрехи“ (ивици на щуки и костури, което ги прави по-малко забележими в гъсталаци на макрофити, ивици на тигри и др.).
Пълна защита от хищници на всички индивиди в популациите на плячки също не се случва, тъй като това би довело не само до смъртта на гладуващите хищници, но в крайна сметка до катастрофа на популациите на плячка. В същото време, при отсъствие или намаляване на гъстотата на популацията на хищниците, генофондът на популацията на плячката се влошава (задържат се болни и стари животни) и поради рязкото увеличаване на броя им се подкопава снабдяването с храна.
Поради тази причина рядко се наблюдава ефектът от зависимостта на размера на популацията на плячката и хищниците - пулсация на размера на популацията на плячката, последвана от пулсация на размера на популацията на хищника с известно закъснение („ефект на Лотка-Волтера“). .
Установява се доста стабилно съотношение между биомасата на хищниците и плячката. Така Р. Риклефс дава данни, че съотношението на биомасата на хищник и жертва варира от 1:150 до 1:300. В различни екосистеми на умерената зона на Съединените щати за един вълк има 300 малки белоопашати елени (тегло 60 кг), 100 големи елени вапити (тегло 300 кг) или 30 лоса (тегло 350). Същият модел е открит в саваните.
При интензивна експлоатация на популациите на фитофагите хората често изключват хищниците от екосистемите (във Великобритания например има сърни и елени, но няма вълци; в изкуствените водоеми, където се отглеждат шарани и други езерни риби, няма щуки). В този случай ролята на хищника се изпълнява от самия човек, премахвайки част от индивидите от популацията на фитофагите.
Особен вариант на хищничество се наблюдава при растенията и гъбите. В растителното царство има около 500 вида, които са способни да хващат насекоми и частично да ги усвояват с помощта на протеолитични ензими. Хищните гъби образуват устройства за улавяне под формата на малки овални или сферични глави, разположени на къси клони на мицела. Най-често срещаният тип капан обаче са адхезивни триизмерни мрежи, състоящи се от голям брой пръстени, образувани в резултат на разклоняването на хифите. Хищните гъби могат да уловят доста големи животни, като кръгли червеи. След като червеят се заплита в хифите, те растат в тялото на животното и бързо го запълват.
1.Постоянни и благоприятни нива на температура и влажност.
2. Изобилие от храна.
3.Защита от неблагоприятни фактори.
4.Агресивен химичен съставместообитание (храносмилателни сокове).
1. Наличието на две местообитания: средата от първи ред е организмът гостоприемник, средата от втори ред е външната среда.
Изв. университети, том 19, 2011 г
ФЕНОМЕН НА МАТЕМАТИЧЕСКИЯ МОДЕЛ НА ЛОТКА-ВОЛТЕРА И ПОДОБНИ НА НЕГО*
DI. Трубецков
Математическият модел на Лотка-Волтера (често наричан модел „хищник-плячка”) е приложим за описание на различни процеси в биологията, екологията, медицината, социалните изследвания, историята, радиофизиката и други науки.
Този преглед, който е до голяма степен методологичен по природа, разглежда варианти на този модел и подобни на него във връзка с анализа на редица природни и социални явления.
Обсъждат се следните модели: модели на взаимодействие между замърсяването и околната среда; модел на класова борба; модел на безкласово общество от ерата на ловците-събирачи; модел на военни действия; вирусен модел на инфекциозно заболяване; модел на разпространение на епидемии, включително модел на заразяване с компютърен вирус; модел на взаимодействието на когнитивните и/или емоционалните режими на мозъка.
Ключови думи: математически модел на Лотка-Волтера, система хищник-жертва, анализ на природни и социални явления.
„...При разрешаване на правни казуси, както при поставяне на диагноза от лекар или по време на експерименти, провеждани от химик, разпознаването на типичните модели играе решаваща роля.“
Бернхард Шлинк. Връщане**
Базиран на въведение екологични примери
В основата си моделът на Лотка-Волтера е математическо описание на дарвиновия принцип на борбата за съществуване, който Чарлз Дарвин излага в автобиографията си по следния начин. „Изследвайки много обстоятелства, стигнах до извода, че има борба за съществуване. Веднъж ми хрумна, че при подобни обстоятелства благоприятните видове ще продължат да се развиват
"Санкт Петербург: Издателска група "ABC-Classics", 2010 г., стр. 67
Пътят, който е следвал Дарвин, е добре описан в книгата. Любопитно е, че Дарвин най-накрая се убеди в заключенията си, след като прочете книгата „Есе върху закона за населението“ на Томас Робърт Малтус. Това есе играе важна роля в социалния и политически живот на Великобритания през 1890 г., тъй като се възприема като разумно разбиране на естествените закони на обществото. През 30-те години малтусианските доктрини потискат правителствените политики. Нека си припомним аргументите на Малтус, които са изненадващо прости. Тъй като, според Малтус, човечеството има естествена тенденция да увеличава своя вид, производството на храна не може да остане на същото ниво.
„Въпреки че съществува приблизителен баланс, както твърди авторът: определен брой хора умират в резултат на природни фактори(глад или болест) или човешки действия (войни, сексуално въздържание или греховни действия като детеубийство), Това е необходимо в името на цялостния баланс на съществуването,
Малтус отиде по-далеч, твърдейки: ... тази съдба (смърт) е участта на най-слабия, най-бедния и най-болния член на обществото себе си, но авторът предупреди за това „Ако дадете милостиня на бедните, човешката раса ще се увеличи и ще има недостиг на храна,“
Викторианските времена сякаш потвърждават, че Малтус е прав. Имаше бунтове за храна, дискусии за закона за бедните и обществена реакция срещу законите за царевицата. През 1834 г. викторианската върхушка, в отговор на поправките към закона за бедните, създава работнически домове, където хората работят за хляб и вода, като алтернатива на църковните общества за благоденствие.
Дарвин е живял в този свят, бил е негов представител и се е движел в същите кръгове като Малтус. Той разпространява идеите на Малтус върху всичко живо: в природата се води борба за живот, борба за съществуване, при която най-лошият и най-слабият организъм умира първи, а по-развитите форми, по-здравите и по-приспособените, побеждават. Именно тези индивиди раждат потомство. Организмите се адаптират по-бързо към условията на средата, ако подобни случаи на борба се повтарят на определени интервали.
Нека започнем по-нататъшното си представяне (като напомняне) с модели на взаимодействащи популации - първите математически модели на Лотка и Волтера. Ще следваме представянето на глава 5 от книгата, тъй като е удобна за по-нататъшно описание на моделите на взаимодействие между замърсяването и околната среда.
Авторите посочват на първо място работата на Волтера, като отбелязват, че подобен модел „беше предложен от Лотка малко по-рано (Волтера извърши много по-пълен анализ на тази система, Лотка всеки път се опитваше да подчертае своя приоритет при записването на този модел) ”,
Моделът на Лотка-Волтера описва взаимодействието на два вида – популация от хищници и популация от плячка.
Нека N(t) е броят на плячката, P(t) е броят на хищниците в момент t. Тогава моделът на Лотка-Волтера има формата:
AN - bNP, - = -dP + cNP, (1)
a, b, c, d са положителни константи.
Нека припомним, че системата от уравнения (1) се основава на следните допускания:
В отсъствието на хищници плячката се възпроизвежда неограничено според уравнението dN/dt = aN, което понякога се нарича уравнение на Малтус;
Хищниците при липса на жертви измират по уравнението dP/dt = -dP;
Членовете, пропорционални на продукта NP, се разглеждат като преобразуване на енергията на един източник в енергията на друг (ефектът от влиянието на популацията на хищници върху популацията на плячката, т.е. резултатът от тяхната среща, е да се намали скоростта на растеж dN/dt на броя на плячката с количеството NP, пропорционално на броя на хищниците).
Ако въведете нови променливи
() N(t)d () P(t)b
n(x) = -v(x) = -^,
тогава системата от уравнения (1) ще приеме формата
N - nv, - = YV(n - 1), dx dx
Системата от уравнения (2) има две фиксирани точки: (0, 0) - седло и (1,1),
за което собствени стойностиЯкобиевите матрици А
- ^1,2 = ±iVУ,
i = l/- 1, т.е. равновесното положение (1.1) е нехиперболично и е невъзможно да се направи заключение за неговата устойчивост въз основа на линеен анализ.
Фазовите траектории на системата от уравнения (2), както е известно, са интегралните криви на уравнението
dv (n - 1)v dn Y (1 - v)n’
чието решение
yn + v - ln nYv = H.
Може да се покаже (вижте например), че в близост до (1, 1) линиите на нивото на функцията H са затворени криви, съответстващи на периодични решения на системата от уравнения (2) (фиг. 1). Константата H в израз (3) се определя от началните условия, например n(0) = n0 и v(0) = vo.
Математическият модел, описан със системата от уравнения (2), е структурен
е нестабилен, т.е. малка промяна в десните части в него може да доведе до качествена промяна в поведението на решението. Това е основният му недостатък. Още повече, че затова е трудно да се говори за съвпадение на теория и експеримент.
Една от причините за структурната нестабилност на модела е, че той е консервативен, т.е. има първи интеграл. Както подчертава авторът на книгата, въпреки посочения основен недостатък на модела, системата от уравнения (1) позволява да се направят нетривиални изводи, потвърдени от многобройни наблюдения. По-специално е формулиран принципът на Волтера: ако в системата „хищник-плячка“, описана от модел (1), двата вида се унищожават равномерно и пропорционално на броя на техните индивиди, тогава средният брой на жертвите се увеличава, а средният броят на хищниците намалява.
Книгата дава два примера за съществуването на този ефект в природата. Единият е социалният факт, довел Волтера до неговата теория за борбата за съществуване: по време на Първата световна война риболовът в Адриатическо море е силно намален, което, за изненада на биолозите, води до увеличаване на хищниците и намаляване на броят на плячката. Вторият пример е по-малко известен. Според принципа на Волтера показва двойнствената природа на използването на репеленти за насекоми (инсектициди) за запазване на културите в полетата. Почти всички подобни химикали действат не само върху вредителите, но и върху техните естествени врагове, което често води до увеличаване на броя на вредителите и намаляване на например птиците, които се хранят с тези вредители. Така, според принципа на Волтера, в екосистема хищник-плячка популацията на плячка е по-чувствителна към процеса на пропорционално намаляване на индивидите в популацията.
В терминологията на V.I. Арнолд, моделът, описан от системата от уравнения (1), се нарича „твърд“. Между другото, част от книгата на V.I. Арнолд „Твърди“ и „меки“ математически модели (M .: MCNO, 2000) се нарича „Твърдите модели като път към погрешни прогнози“. С малка промяна в модела, той става мек, когато се добавят малки членове към дясната страна на (1), като се вземе предвид, например, конкуренцията между плячка за храна или хищници за плячка (дадени са специфични типове уравнения По-долу). След това в общ изгледуравнения (1) могат да бъдат пренаписани, както следва:
В зависимост от малки корекции ef (P, N) и например (P, N), например, сценариите, показани на фиг. 2.
От гледна точка на интерпретацията интересна е фиг. 2, б. Според Арнолд, ако P са бандити, а N са работници, еволюцията на системата води или до рязко увеличаване на броя на бандитите, или до почти пълното им изчезване, тъй като те са ограбили работниците толкова много, че не е останало нищо да взема. Системата в крайна сметка попада в област на стойности на P и N, толкова големи или толкова малки, че моделът става неприложим, защото законите на еволюцията се променят.
Ето няколко по-сложни модела хищник-плячка.
Системата „хищник-плячка“, като се вземе предвид вътреспецифичната конкуренция.
aN - bPN + ef (P, N)
dP + cPN + eg(P,N), напр< 1.
Ориз. 2. а - равновесното състояние е стабилно: за всяка начални условияслед дълго време се установява същото; b - състоянието на равновесие B е нестабилно: системата е „разпространена“; c - в система с нестабилно стационарно състояние A се установява периодичен режим C във времето (за разлика от системата (1) (виж фиг. 1) се извършват колебания с определена амплитуда, независимо от малката отклонение)
където K\,2 са потенциалните възможности на екологичните системи, които се определят от наличното количество ресурси и съответстват на граничните стойности на числеността на населението. В безразмерни променливи тази система от уравнения приема формата
където N(t) = (a/c)u(x), P(t) = av(x)/b, t Конкурентен модел
at, y = d/a, a = a/(cK\), в = d/(aK2).
it1=1-Ш-eNN f=1-f)-hNN (5)
Тук n, r2, e, h са положителни константи.
Анализът на системата от уравнения (5) показва: ако n популации линейно зависят от m ресурси и m< n, то по крайней мере одна из популяций вымирает.
Модел на мутуализъм (симбиоза)
ui(ri + aiiui + ai2u2),
u2 (Г2 + a2iui + a22u2).
За да се опише мутуализмът е достатъчно да се изисква това
ai2 > 0, a2i > 0.
Подробен анализ на горните модели може да се намери например в книгите и.
В края на раздела, след презентацията, разглеждаме обобщения модел на Лотка-Волтера, описан от системата от уравнения
A(u) - B(u,v), - = -D(v) + C (u,v), (7)
където u и v са съответно броят на плячката и хищниците, A(u) е възпроизводствената функция на плячката в отсъствието на хищници; D(v) - функция на изчезване на хищници при липса на плячка; функцията B(u, v) описва изяждането на плячка от хищници; C(u, v) е ефективността на консумация на плячка от хищници. Нека изброим възможните допълнителни фактори на вътрешно- и междупопулационните отношения.
Нелинейна зависимост на скоростта на възпроизводство на популациите на плячка от плътността при ниски стойности на плътност (липса на достатъчен брой чифтосващи двойки): A (u) = au2 / (N + u), където a и N са положителни константи.
Вътрешновидова конкуренция на плячка: A(u) = au(l - u/K).
Наситеност на хищниците: Bi(u) = bu/(1 + au) - трофична функция на хищника.
Нелинеен характер на хищничество от хищници: Bi(u) = bu2/(1 + au).
Конкуренция между хищници за плячка: B2(v) = bv/(1 + |3v). В този случай B(u,v) = Bi(u)B2(v).
Конкуренция на хищници за ресурси, различни от плячката: C(v) = v/(1 + v/Ki).
Нелинеен характер на зависимостта на скоростта на възпроизводство на хищник от плътността на популацията при ниски стойности на плътност: C2(v) = cv/(Nv + v), докато C(u,v) = C1(u)C2(v) и C1(u) = B1(u), D2(v) = cv, т.е. C(u,v) = = cv/(Nv + v) Bi(u).
1. Математически модел на взаимодействието на замърсяването с околната среда, интерпретиран като модел „хищник-плячка“
Ситуацията „замърсяване-природа“ може да се тълкува като специален случаймодел „хищник-плячка“, където природата действа като жертва, а замърсяването – като хищник. Основното предположение, залегнало в модела, е, че околната среда активно абсорбира и обработва замърсяването до определена граница.
От качествени съображения в системата околна среда-замърсяване са възможни следните три фундаментално различни сценария на взаимодействие.
1. С малки емисии на замърсяване околната среда напълно го рециклира (устойчива ситуация).
2. При увеличаване на емисиите на замърсяване, в зависимост от външни условия и случайни причини, околната среда може да бъде в задоволително състояние или да загине (бистабилна ситуация)*.
3. И накрая, третата ситуация съответства на екологична катастрофа - пълното изчезване на природата.
Да приемем, че общият фон на замърсяване и състоянието на околната среда могат да се характеризират със следните променливи: концентрация на замърсяване P и съответно плътност на биомаса E. Ако има постоянен източник на замърсяване, процесът на еволюция на замърсяването може да се опише с уравнението
където a е мощността на източника на замърсяване за единица време, b е коефициентът на естествено унищожаване на замърсяването (такова унищожаване се нарича „мъртво“ или
„Бистабилна ситуация може да възникне, когато в близост до някои предприятия растителната покривка остава само на места. Това следва от наблюдения и в много отношения може да послужи като закъснял сигнал за предприемане на спешни мерки за спасяване на природата.
естествено разсейване). При първоначалното условие P(t = 0) = Po, решението на уравнение (8) има формата:
P (t) = b+(Po - D e~bt’
тоест с течение на времето концентрацията на замърсяване намалява естествено.
Нека освен това приемем, че замърсяването е в постоянно взаимодействие с околната среда, което има почистващ ефект върху замърсяването. Ще обмислим и системата „околна среда – замърсяване” да бъде затворена. Тогава процесът на взаимодействие с околната среда може да се опише със следната система от уравнения:
J- = a - bP- f (E, P), - = g(E) - h(E, P), (9)
където функцията f (E, P) > 0 - описва абсорбцията и обработката на замърсяването на околната среда; g(E) е термин, описващ динамиката на околната среда при липса на замърсяване; h(E, P) е функция, която описва вредното въздействие на замърсяването върху околната среда.
Нека вземем f (E, P) = cEP и h(E, P) = dEP като функции на взаимодействието между замърсяването и дивата природа, където c и d са постоянни коефициенти.
Ще приемем, че при липса на замърсяване поведението на околната среда може да се опише с логистично уравнение, т.е. g(E) = rE(1 - E/K), където r - постоянен коефициент, и K - съответства на максималната стойност на E при dE/dt = 0. Замествайки изразите за f (E, P), h(E, P) и g(E) в системата от уравнения (9), ние добиваме
^ = a - bP-cEP, = rE (l -E) - dEP.
Преминаване в системата от уравнения (10) към безразмерни променливи
T = bt, a = -pr, uo
получаваме най-простия математически модел на взаимодействието на замърсяването с околната среда под формата на следните уравнения:
A - u - uv, dt
v(u0 - u) - pv2.
Лесно се вижда, че системата от уравнения (10) е система хищник-плячка, където плячката е замърсяване (а не биологичен обект), а биологично активната среда е хищникът.
В системата от уравнения (11) параметър а може да се интерпретира като обобщена мощност на източника на замърсяване; uo е максимално допустимата концентрация за дадена екосистема (ако u > u0, dv/dt< 0, и природа вымирает); p - характеристика экосистемы - коэффициент внутривидовой конкуренции природы.
Равновесните положения на системата от уравнения (11), съответстващи на условията du/dt = 0 и dv/dt = 0, се намират лесно от уравненията
a - u(l + v) = 0, -v(u - uo) - pv2 = 0.
Тогава Ai = (a, 0); A2 ((uo + p + Q)/2, (uo - p - Q)/(2p));
A3 ((uo + p - Q)/2, (uo - p + Q)/(2p)), Q = ^/(uo + p)2 - 4ap.
Второто и третото равновесно положение съществуват, ако [(u + p)2-4aP]>0. Стандартният линеен анализ на устойчивостта на посочените равновесни положения води до параметричен портрет на системата от уравнения (11), показана на фиг. 3. На фиг. Фигура 4 показва съответните фазови портрети на системата от уравнения (11).
Ако параметрите принадлежат към област IV, тогава всички траектории с изключение на стабилния колектор Ai се стремят към A3 като t ^ x (фиг. 4, а). Регион II съответства на фазовия портрет на фиг. 4, б. В област III има единичен глобален атрактор A1 (фиг. 4, c).
Така описаното математически моделсъответства на трите качествени ситуации, описани по-горе.
В началото на този раздел обаче беше посочено като основно предположение, че моделът съдържа прагова стойност на замърсяване, която природата може да обработи. Този ефект трябва да се вземе предвид в математическия модел.
Ориз. 3. Параметричен портрет на система от уравнения (11). За фиксирано u0 цялата област на допустимите параметри е разделена на четири групи
За да направите това, книгата предлага да използвате една от трофичните функции, които описват процеса на насищане, например
След това, при ниска концентрация на замърсяване f (E, P) ~ cPE, при висока концентрация f (E, P) cE, което е по-реалистично от оригиналния израз.
След редуциране на системата от уравнения (9) до безразмерна форма, като се вземе предвид връзката (12) и преназначенията в, се получава следната система от уравнения:
A - u ---, - = v(uo - u) - pv , dt k + u dt
където k = A(d/b) > 0 описва степента на влияние на природата върху замърсяването: колкото по-голяма е стойността му, толкова по-малка е степента на усвояване на живата природа и обратно.
В книгата се посочва още, че в системата от уравнения (13) са възможни едно, две или три равновесни състояния от същото естество като за система (11). Освен това авторите подчертават, че априорните допускания относно избора на определени функционални зависимостив този модел са по-адекватни на реалността от избраните модели (11).
2. Математически модел на пречистване на отпадъчни води
Ще разглеждаме замърсителя като „плячка“, а биологично активната утайка като „хищник“. Процесът на биохимично окисляване на замърсител ще се тълкува като неговото „изяждане“ от микроорганизми от активна утайка.
Да приемем, че има постоянен източник на замърсяване и активната утайка е в състояние да преработи замърсяването до определена граница. Смятаме, че промяната в концентрацията на активната утайка в чистата вода намалява по експоненциален закон.
Тогава динамиката на пречистването на отпадъчните води може да се опише с уравненията
dd _ = a - bD(P) - cf (P, E), - = -dS + eh(P, E),
където P (t) е концентрацията на замърсяване на водата, E (t) е плътността на биомасата на активната утайка, D(P) е функцията на разсейване, характеризираща естественото разпадане на замърсяването; f (P, E) и h(P, E) - трофични функции, характеризиращи процеса на пречистване на замърсители с биологично чисти утайки; a > 0 - мощност на източника на замърсяване; d > 0 е константа, характеризираща скоростта на намаляване на активната утайка в чиста вода; c и e са положителни константи.
Да приемем, че D(P) = P, а трофичните функции на „хищника“ и „плячката“ са еднакви и имат вида:
f (P, E) = h(P, E) = , където r > 0 е константа.
Тогава ще имаме:
A - bP--------, -
След въвеждане на безразмерни променливи m = at, P = ru(x), E = (rd/c) v(x), системата от уравнения (14) приема формата
където a = a/(rd), b = b/d, y = e/d. Фиксирани точки на системата (15) Ai(ui,vi) =
= (a/fr 0) и A2(u2, v2) = (1/(Y - 1), Hy - 1) - p]y/(y - 1)).
Стандартният линеен анализ показва, че точката Ai(ui, vi) е седлова точка< a(Y - 1) и y >0 и стабилен възел за β > a(Y - 1).
Ако увеличите стойността на параметъра a при фиксирани стойности в и (Y - 1), което съответства на увеличаване на интензивността на източника на замърсяване, тогава възниква стабилно равновесно състояние A ^ (u2, v2). Ако параметърът a е малък (интензитетът на замърсяване е нисък), тогава масата на биологично активната утайка клони към нула. Тъй като замърсителят е храна за биологично активната утайка, ако интензитетът на замърсителя е недостатъчен, биологично активната утайка изчезва.
Разумно е да се приеме, че има определена прагова стойност на концентрацията на замърсяване P *, над която капацитетът за пречистване на утайките намалява, и да се вземат предвид трофичните функции на вида
f (P, E) = EPe-rP, r = const > 0.
В този случай праговата стойност на замърсяване P* = 1/r. В действителност в описания процес има още един компонент - процесът на аерация - насищане на утайката с кислород, което значително повишава способността на биологично активната утайка да преработва замърсителя. Като се вземе предвид горното, математическият модел на почистване ще приеме формата:
a - bP - cf (P, E),
dE + ef (P, E)Q(t),
където a, b, c, d, e = const > 0. Тук Q(t) е концентрацията на кислород, а R> 0 определя количеството на притока на кислород в системата за единица време.
Анализът на системата от уравнения (16), извършен от авторите на книгата при предположението, че f (P, E) = Pg(E), където g(E) е гладка монотонно нарастваща неотрицателна функция (и g(0) = 0), показва, че при достатъчно близки стойности на параметъра R, който определя количеството кислород, влизащ в системата, винаги е възможно да се постигне стабилност на системата.
3. Моделиране на класовата борба
Нека напишем уравненията на модела "хищник-плячка" под формата на следната система:
където x, y са числата на населението, x1 и y1 са техните стационарни стойности, a и b са константи.
Този модел се използва за описание на малки градски зони. В този случай променливата x в системата от уравнения (17) означава площта на земеползване, y - поземлена рента, a, b, xi, yi - някои параметри.
Интересно приложение на модела за описване на класовата борба е дадено от Гудуин. Нека разгледаме следните два вида граждани: работници и капиталисти.
Работниците изразходват целия си доход wL за потребление, капиталистите натрупват своя доход Y - wL, където Y е производствената продукция. Цената на потребителските стоки се нормализира до единица.
Нека K означава капитал, a = ао egt = Y/L - производителността на труда, нарастваща с постоянен темп d, k = K/Y - коефициентът на капиталоемкост на производството, N = Noent - предлагането на пазара на труда, което нараства с темпа на нарастване на разходите за труд спрямо националния доход е wL/Y = w/a. Следователно делът на печалбата на капиталистите е (1 - w/a). Тъй като спестяването може да се дефинира като S = Y - wL = (1 - w/a)Y, инвестиционният дял е dK/dt = S = (1 - w/a)Y или (dK/dt)(1/K) = (1 - w/a)(Y/K); В този случай пренебрегваме пенсионирането на капитала.
При постоянна стойност на капиталоемкостта k = K/Y получаваме dk/dt = 0 и
dY dL 1 dt Y dt L
В допълнение, от (18), като се вземе предвид факта, че dK/dt = (1 - w/a)L, следва
dY 1 dK 1 / w \ 1
dt Y dt K V a) k-
От съотношения (19) и (20) най-накрая намираме:
Нека въведем нови променливи: y = w/a - дял на разходите за труд; x = L/N - коефициент на заетост.
След прости трансформации, като вземем предвид (21), получаваме
dx - = x dt 1 - y k (d + p)
dt =у w dt a dt
Wf (x), lim f (x) dt
limf(x)< 0,
Линейно приближение на f (x) във формата (dw/dt)(l/w) = -r + bx превръща (23) в следното уравнение:
R + px - g. (24)
Комбинирането на (22) и уравнения
(24), стигаме до модела на Гудуин, описан от системата
y[-(g + r) + px).
Очевидно е, че системата от уравнения (25) е еквивалентна на системата от уравнения (17), следователно всички общи изводи, които са валидни за системата (17), са верни и за модела на Гудуин.
Обърнете внимание, че моделът на Гудуин, който отчита взаимодействието между нивото на заетост и законово установения дял на вноските към заплатите, много напомня на класическите модели на политическата икономия (понякога се нарича неомарксистки). Моделът отново насочва вниманието към трудовете на класическите икономисти като Рикардо, Смит и Маркс. За съжаление моделът е структурно нестабилен.
И така, моделът „хищник“-„плячка“ може да се използва за моделиране на феномените на връзката между рентата за градска земя и интензивността на земеползване, безработицата и динамиката на икономическия растеж.
4. Моделът „хищник-плячка” за описание на безкласово общество – общество от ерата на ловците-събирачи
Тази епоха се характеризира с ниска гъстота на населението и общинна (племенна) организация на човешката общност. Човекът беше напълно зависим от природата, имаше постоянна борба за оцеляване, а основният добиван ресурс беше храната. Човекът беше част от екосистемата и се различаваше от другите училищни хищници по същество само по способността да използва огън и примитивни инструменти за собствените си нужди. Промяната в размера на популацията в определен район се определя главно от състоянието на ресурсната база и може да се опише чрез модела „хищник-плячка“ - система от уравнения (1). Сега в тези уравнения N е размерът на популацията на ловуваните обекти („жертви“); P е броят на древните ловци („хищници“) в разглеждания район; cNP - раждаемост на "хищници"; dP - смъртност на „хищници”; aN е смъртността на „жертвите” (смята се, че хранителните им запаси са неограничени); bNP е степента на смъртност на „плячките“ поради унищожаването им от „хищници“ (счита се, че това е единствената причина за смъртността на „плячките“. Решенията на тези уравнения бяха анализирани във въведението). Тъй като ловците-събирачи консумират произведеното в природата без тяхно участие, размерът на популацията се определя от външни условия, върху които те не могат да повлияят (в модела броят на ловците-събирачи се колебае около средната стойност
Po = a/b. Обърнете внимание, че това обяснява относителната стабилност на населението на Земята през каменната ера до неолита.
Когато човекът опитоми животните и се научи да отглежда култури, той се превърна от ловец-събирач в скотовъдец и земеделец. По този начин коефициентът a и равновесният брой хора се увеличиха значително, тъй като стойността на коефициента b, който характеризира потреблението, остана приблизително на същото ниво.
5. Подобна идеология на хората за описание на военни действия
В книгата Фредерик Ланчестър - английски ученик и инженер - предлага прост модел на борбата на два противника (да речем две армии), който носи неговото име. Имайте предвид, че същият модел е предложен от офицера от царската армия M.O. Осипов през 1915 г. Ето защо е справедливо този модел да се нарече модел на Ланчестър-Осипов. По-нататък в презентацията ще проследим работата. В този модел състоянието на системата се описва от точката (x, y) на положителния квадрант на равнината. Координатите x и y на тази точка са броят на противостоящите армии. Уравненията на модела са както следва:
Чрез, - = -ax. (26)
Тук a и b са мощността на оръжията съответно на армия x и армия y. С други думи, всеки войник от армия x убива войник от армия y за единица време (и, съответно, всеки войник от армия y убива b войника от армия x). По този начин общата ефективност на всички изстрели на армия x се дава от брадва, а за армия y от. Работата предполага, че непрекъснатото приближение е достатъчно добро и скоростта на промяна на числеността на войските е пропорционална на ефективността на изстрелите на противника. „Твърдо“, според V.I. Арнолд, модел (26) допуска точно решение във формата:
ax2 - bу2 = const. (27)
Промяната в броя на армиите x и y става по протежение на хиперболата, дадено от уравнението(27) (фиг. 5). Коя хипербола ще последва войната зависи от началната точка. Хиперболите са разделени от правата линия l/ax = \[bу. Ако началната точка е над тази линия, тогава хиперболата отива към оста y. Това означава, че по време на война размерът на армията x намалява до нула за крайно време. Армията побеждава, врагът е унищожен. Ако началната точка е под линията, армия x печели. По права линия, както пише Арнолд, войната „за всеобщо удовлетворение“ завършва с унищожаването на двете армии. Но това отнема безкрайно дълго време: конфликтът продължава да тлее, когато и двамата опоненти вече са изтощени. От съотношението (27) следва, че на правата ^fax = -\[bу. Тогава първото уравнение на системата (26) приема формата: dx/dt =
Y/abx. Решението му е x = Ce~^a>t, където
С = и е очевидно, че x > 0 при t > Ж Фиг. 5. „Твърд” модел на война
Ориз. 6. „Мек” модел на война
От модел (26) следва, че за борба с двойно по-многоброен враг е необходимо четири пъти по-мощно оръжие, с три пъти повече - девет пъти по-мощно и т.н. Това се показва от квадратните корени в уравнението на правата.
Ясно е, че полученият „канибалистичен“ модел, по терминологията на Арнолд, е силно идеализиран и е опасно да се прилага към реална ситуация. Какво ще се промени, ако
Тази система от уравнения вече не може да бъде решена явно. Но моделът става „мек“ поради промените в a(x, y) и b(x, y). Кривите на равнината (x, y) вече не са хиперболи и не са разделени от права линия (фиг. 6). Но основният качествен извод остава същият: ситуациите „x печели“ и „y печели“ са разделени неутрална линия„двете армии се унищожават една друга за безкраен период от време.“
6. Най-простият вирусен модел на инфекциозно заболяване
През 1974 г. G. Bell предлага най-простия модел на имунния отговор, в който взаимодействието между антиген и антитяло се описва като "хищник-плячка".
Нека представим версия на теорията на G. Bell, базирана на по-общия модел на G.I. Марчук.
Основните фактори в модела на инфекциозното заболяване са следните:
Концентрация на патогенни размножаващи се антигени V (t);
Концентрация на антитела F(t); Антителата означават субстрати на имунната система.
системи, които неутрализират антигени (имуноглобулини, клетъчни рецептори);
Концентрацията на плазмените клетки С - носители и производители на антитела - се приема за постоянна;
Степента на увреждане на целевите органи не се взема предвид.
Уравнението, описващо промяната в броя на антигените (чужди клетки, проникващи в тялото), има формата
W = eV - YFV (28)
Първият член в уравнение (28) е скоростта на растеж на антигените, дължаща се на репродукцията; c - коефициент на размножаване на антигена; терминът yFV описва броя антигени, неутрализирани от F антитела за единица време; y е коефициент, свързан с вероятността за неутрализиране на антигена от антитела при среща с тях.
За да получим второто уравнение, нека изчислим баланса на броя на антителата, които реагират с антигена. Ще има
dF = pCdt - nyFVdt - c/Fdt. (29)
Първият член от дясната страна на уравнение (29) описва генерирането на антитела от плазмените клетки по време на времето dt; p е скоростта на производство на антитяло от плазмените клетки. Вторият термин (nyFVdt) описва намаляването на броя на антителата поради комуникация с антигени и има n антитела за неутрализиране на един антиген. Третият термин (ifFdt) описва намаляването на популацията на антитела поради стареене; if е коефициент, обратно пропорционален на времето на разпадане на антителата. Разделяйки уравнение (29) на dt, получаваме
RS - (ако + nyV)F. (тридесет)
Ако при t< to V(t) = 0, то начальные условия имеют вид
V (до) = Vo, F (до) = Fo. (31)
Нека в това, което следва, let to = 0.
Нека разгледаме два крайни случая на динамика на заболяването. Да приемем, че тялото не произвежда антитела с тази специфичност, тоест F(t) = Fo = 0 за всички t > 0 и p = 0. Тогава от (28) следва, че
- = |3V и V (t) = Voeet, (32)
където Vo е дозата на инфекцията (първоначална концентрация на антигени) при t = 0. Очевидно това решение съответства на хода на заболяването с фатален изход, тъй като няма фактори, компенсиращи растежа на антигените. Вторият ограничаващ случай: наличните в организма антитела (специфични за даден антиген) са достатъчни да унищожат всички антигени, попаднали в тялото, без да се задейства механизмът на образуване на антитела. В този случай в уравнение (28) в ^ yF. Ако приемем, че дозата на инфекцията Vo е малка, стойността на F може да се счита за постоянна, определена от нормалното ниво на антитела F*. Тогава уравнение (28) има формата
F = (v - YF*> V
и неговото решение е написано така:
V = Voe-(YF *-e)t.
В граничния случай, когато B = 0
V = Voe-YF h, (33)
което съответства на висока имунологична бариера.
Решения (32) и (33) са представени на фиг. 7.
За дадени стойности на коефициентите на модела и началните условия е естествено цялото семейство от различни динамики на заболяването да се побере в защрихованата област на фиг. 7.
ограничени от решения за два гранични случая
7. Подобни модели на разпространение на епидемията
Помислете за SIR модела на Kermack-McKendrick, като приемем, че индивидите в една популация могат да бъдат в три различни състояния:
S (t) - здрави индивиди, които са изложени на риск и могат да заразят инфекция;
I(t) - носители на заразени болести;
R(t) - тези, които са се възстановили и са спрели разпространението на болестта (тази категория включва например тези, които са придобили имунитет или дори тези, които са починали).
Типичната еволюция на индивид в популация се описва със следната диаграма:
Нека изградим феноменологичен модел, съответстващ на диаграмата, като използваме следните допускания:
1) популацията е затворена и има постоянен размер N;
2) периодът от време, през който болният остава опасен за другите, се разпределя по експоненциален закон със средно 1/y;
3) контактите са произволни и еднакво възможни (равномерно смесване).
При тези предположения ще имаме:
където X е така наречената сила на инфекцията, която в този случай е равна на произведението от броя на контактите за единица време, направени от един здрав човек (означаваме това число като cN), вероятността от предаване на инфекцията на него при контакт p и вероятността за среща с болен I(t) /N. Нека означим с = cp. Тогава получаваме X = |3! и стигаме до стандартната форма на уравненията:
w dt = S-■<’" ~dt = -о* d + d. + dt = 0и
S (t) + I (t) + R(t) = N.
Третото уравнение в редуцираната система е излишно и тъй като dS/dt< 0, то естественно считать фазовым пространством треугольник S(t) + I(t) < N.
От първото и второто уравнения на системата (34) следва, че
dL =(М-Т^ = -1+ о о = Y I = о dS eSI S’ о в, I =
Като вземем предвид началните условия S(0) = So, I(0) = I0, намираме първия интеграл I + S - o ln S = I0 + S0 - o ln S0, с помощта на който можем да конструираме фаза криви на равнината (S, I) (фиг. 8). Кривите, показани на фиг. 8, b, съвпадат добре с данните за епидемията от чума в Бомбай през 1906 г.
Книгата предлага да се използва моделът, обсъден по-горе, с известна модификация, за анализиране на инфекции с компютърни вируси. Постановката на проблема е следната.
Нека S е броят на компютрите, които са заразени с вирус, I е частта от компютрите, които са заразени с вирус и нямат антивирусен софтуер, а R е частта от компютрите, които имат подходяща антивирусна защита ( имунитет). Тогава уравненията на модела имат формата (вижте системата от уравнения (34)):
^ = -aSI, ^ = aSI - bI, ^ = bI. dt dt dt
Основната идея на модифицирането на този класически епидемичен модел е да се въведе елемент на цикличност, тоест елемент на регулиране на добавянето на нови компютри, податливи на епидемията. Във всеки нов период от време броят на заразените компютри се изчислява въз основа на броя на заразените компютри, останали от предишния момент, а броят на компютрите, изложени на епидемията, се играе като случайна променлива съгласно закона на Поасон с разпределението pm = (Xm/m\)e-x, m = 1,2, ...
Решението на проблема не е дадено. Авторите предлагат следното решение. Постройте графика на функцията I(t) чрез числено решаване на горната система от уравнения с начални условия 5(0) = 100, I(0) = 1, R(0) = 0, X = 10, a = 0,23 , b = 0,5. Предлага се добавяне на нови елементи при tk = k, k = 1, 2, ...100.
Може би някои от читателите ще се заинтересуват от решението на този проблем и многото му възможности.
8. Обобщен модел на Лотка-Волтера за описание
взаимодействия между когнитивните и/или емоционалните режими на мозъка
Според работата широкомащабните когнитивни модели (режими или представяния, наблюдавани в експеримент) в режима на работа на мозъка трябва да се потискат взаимно, което естествено трябва да се случи последователно във времето. С други думи, работещият мозък демонстрира когнитивна и емоционална активност под формата на верига от комбинации от функционални режими, които се сменят един друг с течение на времето, а самите тези комбинации се определят от вида на умствената дейност.
Според авторите „... конкуренцията за ограничени ресурси е една от основните действащи сили, които контролират мозъчните процеси. Тези ресурси са енергия (кислород и глюкоза) и информация (внимание и памет). Важно е информационните ресурси да са продукт на дейността на самия мозък, т.е. те могат да влияят върху емоциите и когнитивните функции по различни начини (например чрез управление на вниманието) и трябва да бъдат включени в общия динамичен модел, който определя работно пространство. ...Режимите, представящи емоциите и когнитивните функции в работното пространство взаимно обменят информация и се конкурират за енергийни и информационни ресурси.“
функционално обусловени от взаимодействието на различни мозъчни подсистеми, тяхната конкуренция и синхронизация (координация) във времето. Горепосочените процеси на конкуренция на когнитивните и емоционалните модуси един с друг, както и емоционалните и когнитивните модуси един с друг, се предлагат да бъдат описани в работата чрез системи от уравнения от типа на Лотка-Волтера в следната обобщена форма: :
Vi(E) -^2 Wi,j(E)xi(t)
Тук Xi(t) характеризира активността на i-тия режим (размера на i-тата популация в екологията); n е броят на взаимодействащите режими (популации); u(E) - информация, постъпваща в системата или налични ресурси; f^- (E) - елементи на матрицата на взаимодействие; n(t) е мултипликативният шум, присъстващ в системата; t е характерното време, което определя процеса.
Ако вземем предвид спецификата на ресурсите, за които се конкурират когнитивните и емоционалните режими, трябва да има три подсистеми от уравнения (36): емоционални режими, когнитивни режими и ресурси (внимание, памет, енергия). Особено важна е ролята на вниманието. В статията за простота се приема ... че описанието на конкуренцията за внимание между емоциите и когнитивните функции не изисква спецификация на отделните режими и следователно можем да се ограничим до разглеждане на конкуренцията за внимание „като цяло ” на емоции B = ^П = 1 Bi и „в общи линии” когнитивни модуси A = ^П=1 Ai. Тогава основните уравнения могат да бъдат записани като:
Oi(I,B,D)Ra -^2 Pij(D)Aj(t)
Zi(S, A,D)Rb -£ lij (D)Bj(t)
Ai(t)n(t), i = 1,...N,
Bi(t)n(t), i = 1, ...M,
RA(t) A - (RA(t) + Fl(1, D)Rb(t)) = Rb(t) B - (Rb(t) + Fb(S, D)RA(t))
В тези уравнения Ai и Bi са неотрицателни променливи, съответстващи на когнитивни и емоционални режими, общата интензивност на които се обозначава съответно с A и B; I - информация от външния свят; S - характеристика на емоционално влияние от външния свят (в случай на отрицателни емоции - стрес); D - ниво на приеманите лекарства; ta и tv са характерни моменти на съзнателна и емоционална дейност; n(t) - мултипликативен шум; коефициентите Fa и Fv определят интензивността на борбата за внимание от страна на емоциите и мисленето. Зависимостта на параметрите Oi и Zi съответно от A и B описва прякото влияние на активността на когнитивните процеси върху емоциите и емоциите върху мисленето. В статията
представен е компютърен анализ на горните уравнения при наличие на външен стимул S(t), характеризиращ нивото на стрес. Обсъжда се и взаимодействието между настроение и мислене. Анализирано е взаимодействието на три компонента на мозъчната дейност: поведение, емоционална памет и когнитивна дейност при липса на външни влияния.
Вместо заключение
В прекрасната книга на Ю.И. Неймарк има такива думи.
„Какво е модел? Думата „модел” е от италиански език и означава „копие, образец, прототип”, изучавайки който се запознаваме с оригинала. Това означава, че математическият модел е прототип на някаква част от света - обект, система, устройство, машина, процес, устройство, чрез изучаването на които ние разбираме тази част от света...”
Ако моделът е стандартен (виж епиграфа) или, както се казва, стандартен, тогава с негова помощ можете да разберете не една, а различни части на света. Математическият модел на Лотка-Волтера е именно такъв модел. Разбира се, представеният преглед обхваща само част от възможните му приложения и не претендира за универсалност.
Библиография
1. гл. Дарвин. Автобиография. 1958. С. 120.
2. Кафява Джанет. Чарлз Дарвин. Произход на видовете / Сер. "10 книги, които промениха света." М.: AST: ast., 2009. 220 с.
3. Малтус Т.Р. Анализ на принципа на населението, тъй като засяга бъдещото подобряване на обществото. 1798 г.
http://www.faculty.rsu.edu/felwell/Theorists/Malthus/essay2.htm
4. Лотка А. Елементи на физическата биология. Балтимор, 1925 г. Препечатано от Dover през 1956 г. като Elements of Mathematical Biology.
5. Волтера В. Математическа теория на борбата за съществуване / Превод. от френски М.: Наука, Главна редакция на физико-математическата литература, 1976. 288 с.
6. Братус А.С., Новожилов А.С., Платонов А.П. Динамични системи и модели на биологията. М.: ФИЗМАТЛИТ, 2010. 400 с.
7. Базикин А.Д. Нелинейна динамика на взаимодействащи популации. Москва-Ижевск: Институт за компютърни изследвания, 2003. 368 с.
8. Арнолд V.I. „Твърди“ и „меки“ модели // Nature. 1998. № 4. С. 3.
9. Братус А.С., Мещерин А.С., Новожилов А.С. Математически модели на взаимодействие между замърсяването и околната среда // Бюлетин на Московския държавен университет. сер. Изчислителна математика и кибернетика. 2001. Т. 6. С. 140.
10. Занг В.-Б. Синергична икономика. Време и промени в нелинейната икономическа теория / Прев. от английски М.: Мир, 1999. 335 с.
11. Гудуин Р.М. Модел на растеж // Социализъм и растеж. Cambridge: University Press, 1967.
12. Малков С.Ю. Социална самоорганизация и исторически процес. Глава 2. М.: URSS, 2009.
13. Lanchester F W. Aircraft in warfire: The down of the fourth arm. Лондон, Констабъл, 1916 г.
14. Осипов М.О. За влиянието на числеността на влизащите в битка сили върху техните загуби // Военен сборник, юни-октомври 1915 г.
15. Bell G. Уравнения за плячка - хищник, симулиращи имунен отговор // Math. Biosci. 1973. № 16. С. 291.
16. Марчук Г.И. Математически модели в имунологията и медицината. Глава 2. М.: Наука, 1985.
17. Kermack W.O. и Mc.Kendrick A.G. Принос към математическата теория на епидемиите // Сборници на Royal Statistical Society A. 1927. Vol. 115. С. 700.
18. Рабинович М.И., Муезинолу М.К. Нелинейна динамика на мозъка: емоции и интелектуална дейност // UFN. 2010. № 4.
19. Неймарк Ю.И. Математически модели в природните науки и технологиите. Н. Новгород: Издателство на университета в Нижни Новгород, 2004 г.
Държава Саратов Получено на 11.02.2011 г
Университет на името на Н.Г. Чернишевски
ФЕНОМЕНЪТ НА МАТЕМАТИЧЕСКИЯ МОДЕЛ НА ЛОТКА-ВОЛТЕРА
И ПОДОБНИ МОДЕЛИ
Математическият модел на Лотка-Волтера (често наричан модел „хищник-плячка”) е приложим за описание на различни процеси в биологията, екологията, медицината, в социологическите изследвания, в историята, радиофизиката и др.
Вариантите на този модел са разгледани методологически в този преглед.
Наблюдават се следните модели: модел на замърсяване или друго мръсно взаимодействие със заобикалящата среда; модел на класова борба; модел на безкласово общество – епоха на ловци-събирачи; модел на военни действия; модел на вирусни инфекциозни заболявания; модел на разпространение на епидемия, така че вирусът на разпространение на компютърна инфекция; модел на когнитивни и (или) емоционални церебрални режими.
Ключови думи: математически модел на Лотка-Волтера, система “хищник-плячка”, анализ на природни или обществени явления.
Трубецков Дмитрий Иванович - роден в Саратов (1938). Завършва Физическия факултет на Саратовския държавен университет (1960 г.). Защитава дисертации за научната степен на кандидат (1965) и доктор на физико-математическите науки в SSU (1978) в областта на радиофизиката. Ръководител на катедрата по електроника, трептения и вълни, Факултет по нелинейни процеси, SSU, професор, член-кореспондент на Руската академия на науките, заслужил учен на Руската федерация, лауреат на Президентската награда за образование. Научен директор на Лицея по приложни науки и Факултета по нелинейни процеси на SSU Област на научни интереси: вакуумна електроника и микровълнова микроелектроника, теория на трептенията и вълните, нелинейна динамика, история на науката. Автор на повече от двадесет учебника и монографии, както и на повече от двеста статии в периодичния печат.
410012, Саратов, ул. Астраханская, 83
Саратовски държавен университет на име. Н.Г. Чернишевски E-mail: [имейл защитен]
Модели на взаимодействие от два типа
Хипотези на Волтера. Аналогии с химичната кинетика. Волтера модели на взаимодействия. Класификация на видовете взаимодействие Конкуренция. Хищник-плячка. Обобщени модели на взаимодействия между видовете . Колмогоров модел. Моделът на Макартър за взаимодействие между два вида насекоми. Параметричен и фазови портрети на системата Базикин.
За основател на съвременната математическа теория на популациите с право се смята италианският математик Вито Волтера, който развива математическата теория на биологичните общности, чийто апарат са диференциални и интегро-диференциални уравнения.(Vito Volterra. Lecons sur la Theorie Mathematique de la Lutte pour la Vie. Париж, 1931). През следващите десетилетия динамиката на популацията се развива главно в съответствие с идеите, изразени в тази книга. Руският превод на книгата на Волтера е публикуван през 1976 г. под заглавието: „Математическа теория на борбата за съществуване“ с послеслов от Ю.М. Свирежев, който разглежда историята на развитието на математическата екология в периода 1931–1976 г.
Книгата на Волтера е написана по начина, по който се пишат книгите по математика. Първо формулира някои предположения относно математическите обекти, които се предполага, че ще бъдат изследвани, и след това провежда математическо изследване на свойствата на тези обекти.
Системите, изследвани от Волтера, се състоят от два или повече типа. В някои случаи се разглежда доставката на използвани храни. Уравненията, описващи взаимодействието на тези типове, се основават на следните концепции.
Хипотези на Волтера
1. Храната или се предлага в неограничени количества, или предлагането й е строго регламентирано във времето.
2. Индивидите от всеки вид умират по такъв начин, че постоянна част от съществуващите индивиди умират за единица време.
3. Хищните видове ядат жертви, като за единица време броят на изядените жертви винаги е пропорционален на вероятността да се срещнат индивиди от тези два вида, т.е. произведението на броя на хищниците и броя на плячката.
4. Ако има храна в ограничени количества и няколко вида, които могат да я консумират, тогава делът на храната, консумирана от даден вид за единица време, е пропорционална на броя на индивидите от този вид, взети с определен коефициент в зависимост от видове (модели на междувидова конкуренция).
5. Ако даден вид се храни с храна, налична в неограничени количества, нарастването на броя на вида за единица време е пропорционално на броя на вида.
6. Ако даден вид се храни с храна, налична в ограничени количества, тогава неговото размножаване се регулира от скоростта на консумация на храна, т.е. за единица време, увеличението е пропорционално на количеството изядена храна.
Аналогии с химичната кинетика
Тези хипотези имат близки паралели с химическата кинетика. В уравненията на динамиката на населението, както и в уравненията на химичната кинетика, се използва "принципът на сблъсъка", когато скоростта на реакцията е пропорционална на произведението на концентрациите на реагиращите компоненти.
Всъщност, според хипотезите на Волтера, скоросттапроцес Изчезването на всеки вид е пропорционално на броя на вида. В химическата кинетика това съответства на мономолекулна реакция на разлагане на определено вещество, а в математическия модел съответства на отрицателни линейни членове от дясната страна на уравненията.
Според концепциите на химическата кинетика, скоростта на бимолекулярната реакция на взаимодействие между две вещества е пропорционална на вероятността от сблъсък на тези вещества, т.е. продукт на тяхната концентрация. По същия начин, в съответствие с хипотезите на Волтера, скоростта на размножаване на хищници (смърт на плячка) е пропорционална на вероятността от срещи между хищници и жертви, т.е. произведението на техните числа. И в двата случая билинейните членове се появяват в моделната система от дясната страна на съответните уравнения.
И накрая, линейните положителни членове от дясната страна на уравненията на Волтера, съответстващи на растежа на популациите при неограничени условия, съответстват на автокаталитичните условия на химичните реакции. Това сходство на уравненията в химичните и екологичните модели ни позволява да прилагаме същите изследователски методи за математическо моделиране на кинетиката на популацията, както за системи от химични реакции.
Класификация на видовете взаимодействия
В съответствие с хипотезите на Волтера, взаимодействието на два вида, числеността на които х 1 и х 2 може да се опише с уравненията:
(9.1)
Ето параметрите а аз - константи на собствената скорост на растеж на вида, ° С аз- константи на самоограничаване на числеността (вътрешновидова конкуренция), b ij- константи за взаимодействие на видовете, (аз, j= 1,2). Знаците на тези коефициенти определят вида на взаимодействието.
В биологичната литература взаимодействията обикновено се класифицират според включените механизми. Разнообразието тук е огромно: различни трофични взаимодействия, химични взаимодействия, съществуващи между бактерии и планктонни водорасли, взаимодействия на гъби с други организми, последователност на растителни организми, свързани по-специално с конкуренцията за слънчева светлина и с еволюцията на почвите и т.н. Тази класификация изглежда обширна.
д . Odum, като взе предвид моделите, предложени от V. Volterra, предложи класификация не по механизми, а по резултати. Според тази класификация връзките трябва да се оценяват като положителни, отрицателни или неутрални в зависимост от това дали изобилието на един вид се увеличава, намалява или остава непроменено в присъствието на друг вид. След това основните видове взаимодействия могат да бъдат представени в табличен вид.
ВИДОВЕ ВЗАИМОДЕЙСТВИЕ НА ВИДОВЕ
|
СИМБИОЗА |
b 12 ,b 21 >0 |
||
|
КОМЕНСАЛИЗЪМ |
b 12 ,>0, b 21 =0 |
||
|
ХИЩНИК-ЖЕРТВА |
b 12 ,>0, b 21 <0 |
||
|
АМЕНЗАЛИЗЪМ |
b 12 ,=0, b 21 <0 |
||
|
КОНКУРЕНЦИЯ |
b 12 , b 21 <0 |
||
|
НЕУТРАЛИЗЪМ |
b 12 , b 21 =0 |
В последната колона са показани знаците на коефициентите на взаимодействие от системата (9.1)
Нека разгледаме основните видове взаимодействия
УРАВНЕНИЯ НА СЪСТЕЗАНИЕТО:
Както видяхме в Лекция 6, уравненията на конкуренцията са:
 (9.2)
(9.2)
Стационарни системни решения:
(1).
![]()
Началото на координатите, за всякакви системни параметри, е нестабилен възел.
(2).
![]() (9.3)
(9.3)
° С стационарно състояние (9.3) е седло при а 1 >б 12 /С 2 и
стабилен възел при а 1 12 /s 2 . Това условие означава, че един вид изчезва, ако неговият собствен темп на растеж е по-малък от определена критична стойност.
(3).
![]() (9.4)
(9.4)
° С стационарно решение (9.4)¾ седло при а 2 >б 21 /° С 1 и стабилен възел при а 2< b 21 /° С 1
(4).
![]() (9.5)
(9.5)
Стационарното състояние (9.5) характеризира съвместното съществуване на два конкуриращи се вида и представлява стабилен възел, ако връзката е изпълнена:
![]()
Това предполага неравенството:
b 12
b 21
което ни позволява да формулираме условието за съвместно съществуване на видовете:
Произведението на коефициентите на междупопулационно взаимодействие е по-малко от произведението на коефициентите на популационното взаимодействие.
Наистина, нека естествените темпове на растеж на двата разглеждани видаа 1 , а 2 са същите. Тогава необходимото условие за стабилност ще бъде
° С 2 > b 12 ,° С 1 >б 21 .
Тези неравенства показват, че увеличаването на размера на един конкурент потиска неговия собствен растеж повече от растежа на друг конкурент. Ако числеността на двата вида е ограничена, частично или напълно, от различни ресурси, горните неравенства са валидни. Ако и двата вида имат абсолютно еднакви нужди, тогава единият от тях ще бъде по-жизнеспособен и ще измести своя конкурент.
Поведението на фазовите траектории на системата дава ясна представа за възможните резултати от конкуренцията. Нека приравним десните части на уравненията на системата (9.2) към нула:
х 1 (а 1 – в 1 х 1 – b 12 х 2) = 0 (dx 1 /dt = 0),
х 2 (а 2 –b 21 х 1 – ° С 2 х 2) = 0 (dx 2 /dt = 0),
В този случай получаваме уравнения за основните изоклини на системата
х 2 = – b 21 х 1 / ° С 2 +а 2 /c 2, х 2 = 0
– уравнения на изоклини на вертикални тангенти.
х 2 = – c 1 х 1 / b 12 + а 1 /б 12 , х 1 = 0
– уравнения на изоклини на вертикални тангенти. Точките на двойно пресичане на изоклините на вертикални и хоризонтални допирателни системи представляват стационарни решения на системата от уравнения (9.2.), а техните координати ![]() са стационарни числа на конкуриращи се видове.
са стационарни числа на конкуриращи се видове.
Възможното разположение на основните изоклини в системата (9.2) е показано на фиг. 9.1. Ориз. 9.1Асъответства на оцеляването на видах 1, фиг. 9.1 b– оцеляване на видах 2, фиг. 9.1 V– съвместно съществуване на видове, когато е изпълнено условие (9.6). Фигура 9.1Ждемонстрира задействащата система. Тук резултатът от състезанието зависи от първоначалните условия. Ненулевото стационарно състояние (9.5) и за двата типа е нестабилно. Това е седлото, през което преминава сепаратриксът, разделяйки зоните на оцеляване на всеки вид.

Ориз. 9.1.Местоположение на главните изоклини на фазовия портрет на системата за конкуренция Волтера от два типа (9.2) с различни съотношения на параметрите. Пояснения в текста.
За да се изследва конкуренцията на видовете, бяха проведени експерименти върху голямо разнообразие от организми. Обикновено се избират два тясно свързани вида, които се отглеждат заедно и поотделно при строго контролирани условия. През определени интервали от време се извършва пълно или избирателно преброяване на населението. Данните от няколко повторени експеримента се записват и анализират. Проведени са изследвания върху протозои (по-специално ресничести), много видове бръмбари от рода Tribolium, дрозофила и сладководни ракообразни (дафния). Проведени са много експерименти върху микробни популации (вижте лекция 11). Проведени са и експерименти в природата, включително върху планарии (Reynolds), два вида мравки (Pontin) и др. На фиг. 9.2. изобразява кривите на растеж на диатомеите, използващи същия ресурс (заемащи същата екологична ниша). При монокултурно отглеждане Asterionella Formosa достига постоянно ниво на плътност и поддържа концентрацията на ресурса (силикат) на постоянно ниско ниво. Б. При отглеждане в монокултураСинедрауина се държи по подобен начин и поддържа концентрацията на силикат на още по-ниско ниво. Б. По време на съвместно култивиране (в два екземпляра) Synedrauina измества Asterionella formosa. Очевидно Синедра

Ориз. 9.2.Конкуренция в диатомеите. А -когато се отглеждат в монокултура Asterionella Formosa достига постоянно ниво на плътност и поддържа концентрацията на ресурса (силикат) на постоянно ниско ниво. б -когато се отглеждат в монокултураСинедрауина се държи по подобен начин и поддържа концентрацията на силикат на още по-ниско ниво. V -със съвместно култивиране (в два екземпляра) Synedruina измества Asterionella formosa. Очевидно Синедра печели конкуренцията поради способността си да използва по-пълно субстрата (вижте също Лекция 11).
Експериментите за изучаване на конкуренцията от G. Gause са широко известни, демонстрирайки оцеляването на един от конкуриращите се видове и му позволявайки да формулира "закона за конкурентно изключване". Законът гласи, че в една екологична ниша може да съществува само един вид. На фиг. 9.3. Резултатите от експериментите на Gause са представени за два вида Parametium, заемащи една и съща екологична ниша (фиг. 9.3 a, b) и видове, заемащи различни екологични ниши (фиг. 9.3 c).
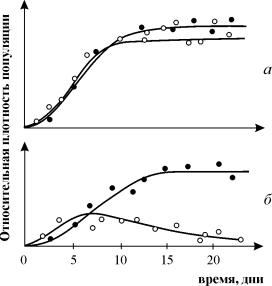
Ориз. 9.3. А- Криви на нарастване на популацията на два видаПараметиум при едновидови култури. Черни кръгове - P Аурелия, бели кръгове – P. Caudatum
b- Криви на растеж на P Aurelia и P . Caudatum в смесена култура.
От Гаузе, 1934 г
Моделът на конкуренцията (9.2) има недостатъци, по-специално следва, че съвместното съществуване на два вида е възможно само ако техният брой е ограничен от различни фактори, но моделът не показва колко големи трябва да бъдат разликите, за да се осигури дългосрочно съвместно съществуване . В същото време е известно, че за дългосрочно съвместно съществуване в променяща се среда е необходима разлика, достигаща определена величина. Въвеждането на стохастични елементи в модела (например въвеждане на функция за използване на ресурси) ни позволява да изследваме количествено тези проблеми.
Система PREDATOR+VICTIM
 (9.7)
(9.7)
Тук, за разлика от (9.2), знаците b 12 И b 21 са различни. Както в случая с конкуренцията, произходът
![]() (9.8)
(9.8)
е специална точка от типа нестабилен възел. Три други възможни стабилни състояния:
![]() ,(9.9)
,(9.9)
![]() (9.10)
(9.10)
![]() (9.11)
(9.11)
Така е възможно да оцелее само плячката (9.10), само хищникът (9.9) (ако има други източници на храна) и съвместното съществуване на двата вида (9.11). Вече разгледахме последния вариант в Лекция 5. Възможните типове фазови портрети за системата хищник-плячка са представени на фиг. 9.4.
Изоклините на хоризонталните допирателни са прави линии
х 2 = – b 21 х 1 /° С 2 + а 1/c 2, х 2 = 0,
и изоклини на вертикални тангенти– прав
х 2 = – ° С 1 х 1 /b 12 + а 2 /b 12 , х 1 = 0.
Стационарните точки се намират в пресечната точка на вертикалната и хоризонталната допирателна изоклина.

От фиг. 9.4 се вижда следното. Система хищник-жертва (9.7) може да има стабилно равновесно положение, в коетоо Ром популацията на жертвите напълно изчезна ( ) и останаха само хищниците (точка 2 на фиг. 9.4 А). Очевидно подобна ситуация може да бъде реализирана само ако освен въпросния тип жертви, х 1 хищник х 2 – има допълнителни източници на захранване. Този факт е отразен в модела чрез положителния член от дясната страна на уравнението за x2. Специални точки(1) и (3) (фиг. 9.4 А) са нестабилни. Втора възможност – стабилно стационарно състояние, при което популацията на хищниците е напълно изчезнала и остава само плячка – стабилна точка(3) (фиг. 9.4 6 ). Тук има специален момент (1) – също нестабилен възел.
И накрая, третата възможност – устойчиво съвместно съществуване на популации от хищник и плячка (фиг. 9.4 V), стационарните числа на които се изразяват с формулите (9.11).
Както в случая с една популация (виж Лекция 3), за модела (9.7) Възможно е да се разработи стохастичен модел, но той не може да бъде решен изрично. Затова ще се ограничим до общи съображения. Да приемем например, че точката на равновесие се намира на определено разстояние от всяка от осите. След това за фазови траектории, на които стойноститех 1 , х 2 остане достатъчно голям, детерминистичният модел ще бъде доста задоволителен. Но ако в даден момент от фазовата траектория някоя променлива не е много голяма, тогава случайните флуктуации могат да станат значителни. Те водят до факта, че изобразяващата точка се премества към една от осите, което означава изчезване на съответния вид.
По този начин стохастичният модел се оказва нестабилен, тъй като стохастичният "дрифт" рано или късно води до изчезване на един от видовете. В този вид модел хищникът в крайна сметка изчезва или случайно, или защото неговата популация на плячка е елиминирана първо. Стохастичният модел на системата хищник-плячка обяснява добре експериментите на Гаузе (Гаузе, 1934), в които ресничките Paramettum candatumслужи като жертва на друга реснички Didinium nasatum – хищник. Очаква се според детерминистични уравнения (9.7) равновесните числа в тези експерименти са били приблизително само пет индивида от всеки вид, така че не е изненадващо, че при всеки повторен експеримент или хищниците, или плячката (и след тях хищниците) измират доста бързо. Резултатите от експериментите са представени на фиг. 9.5.

Ориз. 9.5. Височина Parametium caudatum и хищни реснички Dadinium nasutum. от : Gause G.F. Борбата за съществуване. Балтимор, 1934
И така, анализът на моделите на Volterra за взаимодействие на видове показва, че въпреки голямото разнообразие от типове поведение на такива системи, не може да има незатихващи колебания в числата в модела на конкуриращи се видове изобщо. Такива трептения обаче се наблюдават в природата и в експеримента. Необходимостта от тяхното теоретично обяснение беше една от причините описанието на модела да се формулира в по-обща форма.
Обобщени модели на взаимодействие от два типа
Предложени са голям брой модели за описание на взаимодействието на видовете, чиито десни части на уравненията са функции на броя на взаимодействащите популации. Беше решен въпросът за разработването на общи критерии за установяване какъв тип функции могат да опишат поведението на временния размер на популацията, включително стабилни колебания. Най-известните от тези модели принадлежат на Колмогоров (1935, ревизирана статия - 1972) и Розенцвайг (1963).
 (9.12)
(9.12)
Моделът включва следните допускания:
1) Хищниците не взаимодействат помежду си, т.е. скорост на възпроизвеждане на хищници к 2 и брой жертви Лунищожени за единица време от един хищник не зависи от г.
2) Увеличаването на броя на плячката в присъствието на хищници е равно на увеличението в отсъствието на хищници минус броя на плячката, унищожена от хищници. Функции к 1 (х), к 2 (х), Л(х), - са непрекъснати и определени на положителната полуос х, г³ 0.
3) dk 1 /dx< 0. Това означава, че степента на възпроизводство на плячката в отсъствието на хищник намалява монотонно с увеличаване на броя на плячката, което отразява ограничената наличност на храна и други ресурси.
4) dk 2 /dx> 0, к 2 (0) < 0 < k 2 (¥ ). С увеличаване на броя на жертвите, коефициентът на възпроизводство на хищниците намалява монотонно с увеличаване на броя на жертвите, преминавайки от отрицателни стойности (когато няма какво да се яде) към положителни.
5) Броят на плячката, унищожена от един хищник за единица време Л(х)> 0 при N> 0; Л(0)=0.
Възможните видове фазови портрети на системата (9.12) са представени на фиг. 9.6:

Ориз. 9.6.Фазови портрети на системата на Колмогоров (9.12), която описва взаимодействието на два типа при различни съотношения на параметрите. Пояснения в текста.
Стационарните решения (има две или три) имат следните координати:
(1). ` х=0;` y=0.
Произходът на координатите за всякакви стойности на параметрите е седло (фиг. 9.6 a-d).
(2). ` x=A,` y=0.(9.13)
Аопределя се от уравнението:
к 1 (А)=0.
Стационарен решение (9.13) е седло, ако б< А (фиг. 9.6 А, b, Ж), б определен от уравнението
к 2 (б)=0
Точка (9.13) се поставя в положителния квадрант, ако B>A . Това е стабилен възел .
Последният случай, който съответства на смъртта на хищника и оцеляването на плячката, е показан на фиг. 9.6 V.
(3). ` x=B,` y=C.(9.14)
Стойността на C се определя от уравненията:
Точка (9.14) – фокус (фиг.9.6 А) или възел (фиг.9.6 Ж), чиято стабилност зависи от знака на количествотос
с 2 = – к 1 (B) – k 1 (б)B+L(б)° С.
Ако с>0, точка е стабилна, акос<0 ‑ точка неустойчива, и вокруг нее могут существовать предельные циклы (рис. 9.6 b)
В чуждестранната литература по-често се разглежда подобен модел, предложен от Rosenzweig и MacArthur (1963):
 (9.15)
(9.15)
Където f(х) - скорост на изменение на броя на жертвите хв отсъствието на хищници, F( x,y) - интензивност на хищничество, к- коефициент, характеризиращ ефективността на преработката на биомаса от плячка в биомаса от хищници, д- смъртност на хищници.
Модел (9.15) се свежда до специален случай на модела на Колмогоров (9.12) при следните допускания:
1) броят на хищниците е ограничен само от броя на плячката,
2) скоростта, с която даден хищник изяжда плячка зависи само от плътността на популацията на плячката и не зависи от плътността на популацията на хищника.
Тогава уравненията (9.15) приемат формата.

Когато се описва взаимодействието на реални видове, десните страни на уравненията се уточняват в съответствие с идеите за биологичните реалности. Нека разгледаме един от най-популярните модели от този тип.
Модел на взаимодействие между два вида насекоми (Макартър, 1971 г.)
Моделът, който ще разгледаме по-долу, беше използван за решаване на практическия проблем за борба с вредните насекоми чрез стерилизация на мъжките от един от видовете. Въз основа на биологичните особености на взаимодействието на видовете е написан следният модел
 (9.16)
(9.16)
Тук x,y- биомаса от два вида насекоми. Трофичните взаимодействия на видовете, описани в този модел, са много сложни. Това определя формата на полиномите от дясната страна на уравненията.
Нека да разгледаме дясната страна на първото уравнение. Видове насекоми хядат ларвите на вида при(член +k 3 y),но възрастните от вида приядат ларвите на вида хобект на голямо изобилие на видове хили приили и двата типа (членове – к 4 xy, – y 2). На малки хвидова смъртност хпо-висок от естествения му прираст (1 –к 1 +k 2 x–x 2 < 0 на малки Х).Във второто уравнение терминът к 5 отразява естествения растеж на вида y; –к 6 y –самоограничение от този тип,–к 7 х– изяждане на ларви от вида привидове насекоми х, к 8 xy – увеличаване на биомасата на вида припоради консумация от възрастни насекоми от вида приларви на вида Х.
На фиг. 9.7 представен е граничен цикъл, който е траекторията на устойчиво периодично решение на системата (9.16).

Решението на въпроса как да се осигури съвместното съществуване на населението с неговата биологична среда, разбира се, не може да бъде получено без отчитане на спецификата на дадена биологична система и анализ на всички нейни взаимовръзки. В същото време изследването на формалните математически модели ни позволява да отговорим на някои общи въпроси. Може да се твърди, че за модели като (9.12) фактът на съвместимост или несъвместимост на популациите не зависи от първоначалния им размер, а се определя само от естеството на взаимодействието на видовете. Моделът помага да се отговори на въпроса: как да повлияем на биоценозата и да я управляваме, за да унищожим бързо вредните видове.
Управлението може да се сведе до краткосрочна, рязка промяна в стойностите на населението хИ u.Този метод съответства на методи за контрол като еднократно унищожаване на една или и на двете популации чрез химически средства. От формулираното по-горе твърдение става ясно, че за съвместими популации този метод на контрол ще бъде неефективен, тъй като с времето системата отново ще достигне стационарен режим.
Друг начин е да промените типа на функциите за взаимодействие между изгледите, например, когато променяте стойностите на системните параметри. Именно на този параметричен метод отговарят методите за биологичен контрол. По този начин, когато се въвеждат стерилизирани мъжки, скоростта на естествения прираст на популацията намалява. Ако в същото време получим различен тип фазов портрет, при който има само стабилно стационарно състояние с нулев брой вредители, контролът ще доведе до желания резултат – унищожаване на популацията на вреден вид. Интересно е да се отбележи, че понякога е препоръчително въздействието да се прилага не върху самия вредител, а върху неговия партньор. Като цяло е невъзможно да се каже кой метод е по-ефективен. Това зависи от наличните контроли и от изричната форма на функциите, описващи взаимодействието на популациите.
Модел на A.D. Базикин
Теоретичният анализ на моделите на взаимодействието на видовете е най-изчерпателно извършен в книгата на A.D. Bazykin "Биофизика на взаимодействащите популации" (М., Наука, 1985).
Нека разгледаме един от моделите хищник-плячка, изучавани в тази книга.
 (9.17)
(9.17)
Система (9.17) е обобщение на най-простия модел Volterra хищник-плячка (5.17), като взема предвид ефекта от насищането на хищник. Модел (5.17) предполага, че интензивността на пашата на плячката нараства линейно с увеличаване на плътността на плячката, което не отговаря на реалността при висока плътност на плячката. Могат да бъдат избрани различни функции, за да се опише зависимостта на диетата на хищника от плътността на плячката. Най-важно е избраната функция да расте хклони асимптотично към постоянна стойност. Модел (9.6) използва логистична зависимост. В модела на Базикин хиперболата е избрана като такава функция х/(1+px). Нека припомним, че това е формата на формулата на Моно, която описва зависимостта на скоростта на растеж на микроорганизмите от концентрацията на субстрата. Тук плячката действа като субстрат, а хищникът действа като микроорганизми. .
Системата (9.17) зависи от седем параметъра. Броят на параметрите може да бъде намален чрез замяна на променливи:
х® (A/D)х; г ® (A/D)/y;
T® (1/А)T; g (9,18)
и зависи от четири параметъра.
За цялостно качествено изследване е необходимо четиримерното параметрично пространство да се раздели на области с различни видове динамично поведение, т.е. изградете параметричен или структурен портрет на системата.
След това е необходимо да се построят фазови портрети за всяка от областите на параметричния портрет и да се опишат бифуркациите, които възникват с фазовите портрети на границите на различните области на параметричния портрет.
Изграждането на пълен параметричен портрет се извършва под формата на набор от „резове“ (проекции) на нискоразмерен параметричен портрет с фиксирани стойности на някои от параметрите.
Параметричен портрет на системата (9.18) за фиксиран жи малки дпредставени на фиг. 9.8. Портретът съдържа 10 области с различни типове поведение на фазовите траектории.
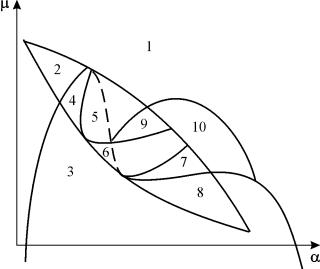
Ориз. 9.8.Параметричен портрет на системата (9.18) за фиксиранж
и малки д
Поведението на системата при различни съотношения на параметрите може да бъде значително различно (фиг. 9.9). Системата позволява:
1) едно стабилно равновесие (региони 1 и 5);
2) един стабилен граничен цикъл (региони 3 и 8);
3) две стабилни равновесия (регион 2)
4) стабилен граничен цикъл и нестабилно равновесие вътре в него (региони 6, 7, 9, 10)
5) стабилен граничен цикъл и стабилно равновесие извън него (област 4).
В параметричните области 7, 9, 10 областта на привличане на равновесие е ограничена от нестабилен граничен цикъл, лежащ вътре в стабилен. Най-интересната структура е фазовият портрет, съответстващ на зона 6 в параметричния портрет. Подробно е показано на фиг. 9.10.

Зоната на привличане на равновесие B 2 (защрихована) е „охлюв“, усукващ се от нестабилния фокус B 1. Ако е известно, че в началния момент от време системата е била в близост до B 1, тогава е възможно да се прецени дали съответната траектория ще достигне равновесието B 2 или стабилен граничен цикъл около трите равновесни точки C (седло ), B 1 и B 2 въз основа на вероятностни съображения.

Фиг.9.10.Фазов портрет на система 9.18 за параметрична област 6. Област на привличане B 2 е защрихована
В параметричен портрет(9.7) има 22 различни бифуркационни граници, които образуват 7 различни видове бифуркации. Тяхното изследване ни позволява да идентифицираме възможните типове поведение на системата, когато нейните параметри се променят. Например при движение от района 1 към зона 3 възниква раждането на малък граничен цикъл или мекото раждане на автоколебания около едно равновесие IN.Подобно меко раждане на собствени трептения, но около едно от равновесните положения, а именно б 1 , възниква при преминаване на границите на регионите 2 и 4. При напускане на района 4 към зона 5 стабилен граничен цикъл около точкаб 1 “избухва” на примката от сепаратриси и единствената привличаща точка остава равновесието б 2 и т.н.
От особен интерес за практиката е, разбира се, разработването на критерии за близостта на системата до бифуркационните граници. Наистина, биолозите са добре запознати със свойството „буфериране“ или „гъвкавост“ на естествените екологични системи. Тези термини обикновено се отнасят до способността на системата да абсорбира външни влияния. Докато интензивността на външното въздействие не надвишава определена критична стойност, поведението на системата не претърпява качествени промени. На фазовата равнина това съответства на връщането на системата към стабилно състояние на равновесие или към стабилен граничен цикъл, чиито параметри не се различават много от първоначалния. Когато интензитетът на въздействието надхвърли допустимото, системата се „разваля” и преминава в качествено различен режим на динамично поведение, например просто умира. Това явление съответства на бифуркационен преход.
Всеки тип бифуркационен преход има свои отличителни черти, които позволяват да се прецени опасността от такъв преход за екосистемата. Ето някои общи критерии, показващи близостта на опасна граница. Както в случая с един вид, ако при намаляване на числеността на един от видовете системата „заседне“ в близост до нестабилна седлова точка, което се изразява в много бавно възстановяване на числеността до първоначалната стойност, тогава системата е близо до критичната граница. Показател за опасност е и промяната във формата на колебанията в числеността на хищниците и жертвите. Ако колебанията, близки до хармоничните, се превърнат в релаксационни и амплитудата на колебанията се увеличи, това може да доведе до загуба на стабилност на системата и изчезване на един от видовете.
По-нататъшното задълбочаване на математическата теория за взаимодействието между видовете върви по линията на детайлизиране на структурата на самите популации и отчитане на времеви и пространствени фактори.
Литература.
Колмогоров A.N. Качествено изследване на математически модели на динамиката на населението. // Проблеми на кибернетиката. М., 1972, бр.5.
MacArtur R. Графичен анализ на екологичните системи // Доклад на отдела по биология Perinceton University. 1971
A.D. Базикин „Биофизика на взаимодействащите популации“. М., Наука, 1985.
В. Волтера: „Математическа теория за борбата за съществуване“. М.. Наука, 1976
Gause G.F. Борбата за съществуване. Балтимор, 1934 г.
Динамиката на населението е един от клоновете на математическото моделиране. Интересен е, защото има специфични приложения в биологията, екологията, демографията и икономиката. В този раздел има няколко основни модела, един от които, моделът „Хищник-Плячка“, се обсъжда в тази статия.
Първият пример за модел в математическата екология е моделът, предложен от V. Volterra. Той беше този, който за първи път разгледа модела на връзката между хищник и плячка.
Нека разгледаме постановката на проблема. Нека има два вида животни, едното от които поглъща другото (хищници и плячка). В този случай се правят следните предположения: хранителните ресурси на плячката не са ограничени и следователно, при липса на хищник, популацията на плячката нараства по експоненциален закон, докато хищниците, отделени от жертвите си, постепенно умират на глада, също според експоненциален закон. След като хищниците и плячката започнат да живеят в непосредствена близост един до друг, промените в размера на популацията им стават взаимосвързани. В този случай очевидно относителното увеличение на броя на плячката ще зависи от размера на популацията на хищниците и обратно.
В този модел се приема, че всички хищници (и цялата плячка) са в едни и същи условия. В същото време хранителните ресурси на жертвите са неограничени, а хищниците се хранят изключително с жертвите. И двете популации живеят в ограничен район и не взаимодействат с други популации и няма други фактори, които биха могли да повлияят на размера на популацията.
Самият математически модел „хищник-плячка“ се състои от двойка диференциални уравнения, които описват динамиката на популациите от хищници и плячка в най-простия случай, когато има една популация от хищници и една от плячка. Моделът се характеризира с колебания в размера на двете популации, като пикът при хищниците е малко зад пика при плячката. Този модел може да се намери в много трудове по динамика на населението или математическо моделиране. Той е широко застъпен и анализиран с помощта на математически методи. Формулите обаче не винаги могат да предоставят очевидна представа за протичащия процес.
Интересно е да разберете как точно динамиката на популацията в този модел зависи от първоначалните параметри и доколко това отговаря на реалността и здравия разум и да видите това графично, без да прибягвате до сложни изчисления. За целта по модела Volterra е създадена програма в среда Mathcad14.
Първо, нека проверим модела за съответствие с реалните условия. За да направим това, нека разгледаме изродени случаи, когато само една от популациите живее при дадени условия. Теоретично е доказано, че при отсъствие на хищници популацията на плячката се увеличава неограничено във времето, а популацията на хищник при липса на плячка измира, което като цяло отговаря на модела и реалната ситуация (с посочената формулировка на проблем).
Получените резултати отразяват теоретичните: хищниците постепенно измират (фиг. 1), а броят на плячката се увеличава неограничено (фиг. 2).
Фиг. 1 Зависимост на броя на хищниците от времето при липса на плячка

Фиг. 2 Зависимост на броя на плячката от времето при липса на хищници
Както се вижда, в тези случаи системата съответства на математическия модел.
Нека разгледаме как се държи системата при различни начални параметри. Нека има две популации - лъвове и антилопи - съответно хищници и плячка и са дадени начални показатели. Тогава получаваме следните резултати (фиг. 3):
Таблица 1. Коефициенти на колебателен режим на системата

Фиг.3 Система със стойности на параметрите от Таблица 1
Нека анализираме данните, получени въз основа на графиките. С първоначалното увеличаване на популацията на антилопите се наблюдава нарастване на броя на хищниците. Обърнете внимание, че пиковото увеличение на популацията на хищници се наблюдава по-късно, по време на намаляването на популацията на плячката, което е напълно в съответствие с реалните концепции и математическия модел. Всъщност увеличаването на броя на антилопите означава увеличаване на хранителните ресурси за лъвовете, което води до увеличаване на техния брой. Освен това, активната консумация на антилопи от лъвове води до бързо намаляване на броя на плячката, което не е изненадващо, като се има предвид апетита на хищника или по-скоро честотата на хищниците, които ядат плячка. Постепенното намаляване на броя на хищниците води до ситуация, при която популацията на плячката се оказва в условия, благоприятни за растеж. След това ситуацията се повтаря с определен период. Ние заключаваме, че тези условия не са подходящи за хармоничното развитие на индивидите, тъй като те водят до рязко намаляване на популацията на плячката и рязко увеличаване на двете популации.
Нека сега зададем началната численост на хищника, равна на 200 индивида, като запазим други параметри (фиг. 4).
Таблица 2. Коефициенти на колебателен режим на системата

Фиг.4 Система със стойности на параметрите от Таблица 2
Сега системата осцилира по-естествено. При тези предположения системата съществува доста хармонично, няма резки увеличения и намаления на числеността и в двете популации. Заключаваме, че с тези параметри и двете популации се развиват достатъчно равномерно, за да живеят заедно на една и съща територия.
Нека зададем първоначалния брой на хищниците на 100 индивида, броя на плячката на 200, като запазим други параметри (фиг. 5).
Таблица 3. Коефициенти на колебателен режим на системата

Фиг.5 Система със стойности на параметрите от Таблица 3
В този случай ситуацията е близка до първата разгледана ситуация. Обърнете внимание, че при взаимно увеличаване на популациите, преходите от увеличаване към намаляване на популацията на плячка стават по-плавни и популацията на хищници остава при липса на плячка на по-висока числена стойност. Ние заключаваме, че когато една популация е тясно свързана с друга, тяхното взаимодействие се осъществява по-хармонично, ако специфичните първоначални популации са достатъчно големи.
Нека разгледаме промяната на други системни параметри. Нека началните числа отговарят на втория случай. Да увеличим скоростта на възпроизводство на жертвите (фиг. 6).
Таблица 4. Коефициенти на колебателен режим на системата

Фиг.6 Система със стойности на параметрите от Таблица 4
Нека сравним този резултат с резултата, получен във втория случай. В този случай се наблюдава по-бърз растеж на жертвата. В този случай както хищникът, така и жертвата се държат като в първия случай, което се обяснява с ниския размер на популацията. При това взаимодействие и двете популации достигат връх при стойности, много по-големи, отколкото във втория случай.
Сега нека увеличим скоростта на растеж на хищниците (фиг. 7).
Таблица 5. Коефициенти на колебателен режим на системата

Фиг.7 Система със стойности на параметрите от таблица 5
Нека сравним резултатите по подобен начин. В този случай общите характеристики на системата остават същите, с изключение на промяната в периода. Както може да се очаква, периодът се скъси, което се обяснява с бързото намаляване на популацията на хищниците при липса на плячка.
И накрая, нека променим коефициента на междувидово взаимодействие. Първо, нека увеличим честотата на хищници, които ядат плячка:
Таблица 6. Коефициенти на колебателен режим на системата

Фиг.8 Система със стойности на параметрите от Таблица 6
Тъй като хищникът яде плячката си по-често, максималният размер на популацията се е увеличил в сравнение с втория случай, а разликата между максималния и минималния размер на популацията също е намаляла. Периодът на трептене на системата остава същият.
А сега нека намалим честотата на яденето на плячка от хищници:
Таблица 7. Коефициенти на колебателен режим на системата

Фиг.9 Система със стойности на параметрите от Таблица 7
Сега хищникът яде плячката по-рядко, максималният размер на популацията е намалял в сравнение с втория случай, а максималният размер на популацията на плячката се е увеличил 10 пъти. От това следва, че при тези условия популацията на плячката има по-голяма свобода по отношение на възпроизводството, тъй като хищникът се нуждае от по-малко маса, за да се насити с нея. Разликата между максималния и минималния размер на популацията също е намаляла.
Когато се опитваме да моделираме сложни процеси в природата или обществото, по един или друг начин възниква въпросът за коректността на модела. Естествено, при моделирането процесът се опростява и някои незначителни детайли се пренебрегват. От друга страна, съществува опасност моделът да се опрости твърде много, като по този начин се изхвърлят важни характеристики на явлението заедно с маловажните. За да се избегне тази ситуация, преди моделирането е необходимо да се проучи предметната област, в която се използва този модел, да се проучат всички негови характеристики и параметри и най-важното, да се подчертаят тези характеристики, които са най-значими. Процесът трябва да има естествено описание, интуитивно разбираемо, съвпадащо в основните моменти с теоретичния модел.
Моделът, разгледан в тази работа, има редица съществени недостатъци. Например допускането на неограничени ресурси за жертвата, липсата на фактори от трети страни, влияещи върху смъртността и на двата вида и т.н. Всички тези предположения не отразяват реалната ситуация. Но въпреки всички недостатъци, моделът е широко разпространен в много области, дори далеч от екологията. Това може да се обясни с факта, че системата "хищник-плячка" дава обща представа за взаимодействието на видовете. Взаимодействията с околната среда и други фактори могат да бъдат описани с други модели и анализирани заедно.
Отношенията от типа "хищник-жертва" са съществена характеристика на различни видове жизнена дейност, в които има сблъсък между две взаимодействащи страни. Този модел има място не само в екологията, но и в икономиката, политиката и други области на дейност. Например една от областите, свързани с икономиката, е анализът на пазара на труда, като се вземат предвид наличните потенциални работници и свободните работни места. Тази тема би била интересно продължение на работата по модела хищник-плячка.
Нека и са съответно броят на плячката и хищниците. Нека приемем, че единственият ограничаващ фактор, ограничаващ размножаването на плячката, е натискът върху тях от хищници, а размножаването на хищниците е ограничено от количеството храна, която получават (броя на плячката). Тогава, при липса на хищници, броят на плячката трябва да нараства експоненциално с относителна скорост a, а хищниците при липса на плячка също трябва да измират експоненциално с относителна скорост t плячка и съответно естествена смъртност на хищниците.
Нека е броят (или биомасата) на плячката, консумирана от един хищник за единица време, като част от енергията, получена от тази биомаса, се изразходва от хищника за възпроизвеждане, а останалата част се изразходва за поддържане на основния метаболизъм и ловна дейност. Тогава уравнението на системата хищник-плячка може да бъде написано като

Функцията обикновено се нарича трофична функция на хищника или функционален отговор (функционален отговор) на хищника към гъстотата на популацията на плячката. Именно тези функции обикновено се идентифицират в експерименталните изследвания на хищничеството и сега се приема, че тези функции обикновено принадлежат към един от следните три типа (фиг. 11). Очевидно динамичното поведение на системата до голяма степен зависи от вида на трофичната функция.

Ориз. 11. Различни видове трофични функции в системата хищник-жертва: а) този тип е характерен за безгръбначните и някои видове хищни риби; б) трофична функция с ясно изразен праг на насищане е характерна за хищниците, хранещи се с филтър (например много мекотели); в) този тип е характерен за гръбначните животни - организми, които показват доста сложно поведение (например способни да учат). Трофичната функция ще има подобен вид, ако плячката може да развие отбранителна стратегия (например скриване в убежище, недостъпно за хищници).
За малки стойности на x, например, когато трофичните отношения в системата са напрегнати и почти цялата плячка става плячка на хищник, който винаги е гладен и не се засища (ситуацията е доста често срещана в природата), трофичната функция може да се счита за линейна функция на броя на плячката, т.е. в допълнение, нека приемем, че . Тогава

Система (2.2) съвпада до нотация с класическия модел хищник-плячка на V. Volterra, който показа, че тази система има интеграл от формата
![]()
където If са първоначалните стойности на броя на плячката и съответно хищниците, тогава
![]()
и уравнение (2.3) описва семейство от затворени криви, вложени една в друга, съответстващи на фазовите траектории на периодични решения на система (2.2).

Ориз. 12. Фазов портрет на класическата система Волтера хищник - плячка.
Обърнете внимание, че с увеличаването на C амплитудите на трептенията и y се увеличават (фиг. 12). При минималната стойност тези криви се свиват до точка с координати . Лесно се вижда, че те са решения на система (2.2) за , т.е. нейното нетривиално равновесие. За случая на малки колебания близо до това състояние уравненията на модела могат да бъдат записани като
![]()
Корените на съответното характеристично уравнение са , т.е. точката е центърът. Период на малки колебания, като колебанията в броя на един вид се изместват във фаза спрямо колебанията на друг с
В системата (2.2) има още едно равновесно положение - началото. Лесно се вижда, че тази точка е седло. Координатните оси са сепаратриси, като оста влиза в седлото, а оста го напуска (виж фиг. 12).
Въпреки факта, че моделът на Волтера успя да обясни много реално наблюдавани явления, той има голям недостатък - негрубостта (в математическия смисъл на думата) на циклите на Волтера, така че за всякакви, колкото и слаби, смущения на фазовите координати, системата преминава от един цикъл към друг. Очевидно по-адекватните модели трябва да имат това свойство на „грапавост“.
От гледна точка на теорията на устойчивостта, равновесното състояние на системата (x, y) е състояние на безразлично равновесие, стабилно според Ляпунов, но не асимптотично липсата на асимптотична стабилност на равновесието показва, че в системата Волтера няма механизми, които да се стремят да запазят неговото нетривиално равновесно състояние.
Горното твърдение за стабилността на равновесието се доказва доста лесно чрез конструиране на съответната функция на Ляпунов. Но повече за това в следващия параграф.

Подобни статии



